Origins of Life
Freeman DYSON
Origine de la vie : n'oublions pas l'azote
Mise à jour régulière d'un texte publié dans le "Dictionnaire de l'Ignorance" Presses Universitaires de France, Paris, (1998) pp. 196-212 (© 2000 Antoine Danchin DR) et d'une conférence au Collège de France à l'invitation de Jacques Roger.
Noter qu'un texte en anglais développe un certain nombre de points qui ne sont pas traités ici. En particulier, les contraintes physico-chimiques de la matière limitent très fortement les possibilités de création de macromolécules informationnelles. Ce n'est donc qu'à la suite d'expériences mal conduites, relayées par une grave distorsion de ce que devraient être les publications scientifiques, et d'une pression médiatique ignorante qu'on a pu croire que l'arsenic pourrait faire partie du squelette des acides nucléiques. Tout au plus pourrait-il intervenir comme "décoration" de ce squelette, mais certainement pas pour remplacer la liaison phosphodiester.
La réflexion sur le métabolisme de surface doit à de nombreux auteurs, mais nous devons retenir en priorité le raisonnement de Freeman Dyson qui a montré très explicitement que la reproduction (faire une copie semblable) devait précéder la réplication (faire une copie exacte). Le métabolisme est donc antérieur au monde des ARN, qui, lui-même doit se diviser en une première étape, métabolisme général, synthèse des nucléotides et monde des ARN, et une deuxième étape, où apparaît la réplication, le monde des génomes à ARN. Il est aussi essentiel de comprendre que, du point de vue de la chimie, il existe une logique du métabolisme, plutôt qu'un assemblage incompréhensible de réactions chimiques que les biochimistes apprennent par cœur.
Un scénario original, peu à peu élaboré à partir du début des années 2000, propose de reconsidérer l'origine des premières cellules à partir de la distinction faite par Dyson, pour imaginer un ensemble d'assez grosses cellules capables de fusionner ou de s'ingérer les unes les autres. Au sein de ces cellules deux compartiments se développent, le compartiment de l'ARN métabolisme où les ancêtres de ribosomes vont synthétiser les protéines actives, et le compartement de l'ARN génome, ancêtre du noyau, qui conserve en mémoire le pnan de construction des protéines et des ribosomes. Cette situation instable va se pérenniser grâce à la découverte de l'ADN, en plusieurs étapes et en plusieurs endroits. Il n'y a pas d'unique ancêtre des cellules, mais une collection de cellules différentes qui fusionnent et échangent leurs innovations. Ces cellules prédatrices vont voir apparaître un mode de résistance à la prédation sous la forme de deux classes de plus petites cellules, qui seront les bactéries et les archées. Ce scénario est élaboré dans deux arcticles de Médecine/Sciences publiés à la fin de l'année 2018.
Retour sur les
origines de la vie:
De
l'atome aux molécules, reproduction réplication
Des
métabolites aux cellules
Pour des compléments et des détails des processus nécessaires à la création de la vie, voir aussi : Texte de la conférence donnée à Rio de Janeiro pour le 100ème anniversaire de la mort de Louis Pasteur (en anglais, Février 1995), présentation de la conférence de Rio, une conférence au Centre International de Synthèse, et (Le Seuil 1990) (résumé). Il est important aussi de comprendre que la comparaison des génomes, de façon extraordinaire, nous donne accès à une archive de l'origine de la vie (résumé en anglais), et nous guide vers ce qui a pu se produire il y a près de quatre milliards d'années.
Conférence à l'Académie des Sciences, 16 septembre 2013 : l'invention des premières cellules
Cristallographe, Louis
Pasteur fit l'étonnante découverte d'un caractère original qui
allait de pair avec la vie : les molécules qui dérivent des processus
vivants cristallisent sous une forme dissymétrique. Alors que les
molécules issues de la chimie ordinaire donnent souvent un mélange de
cristaux symétriques les uns des autres comme le sont les mains, ou
les objets avec leur reflet dans un miroir, le tartrate présent dans
la lie du vin ne donnait qu'un seul type de cristal, sans qu'on puisse
y découvrir son symétrique. Il a donc paru nécessaire à Pasteur
d'inclure un processus spécifique dans la chimie du vivant afin de la
différencier de la chimie ordinaire. En raison de ses convictions
philosophiques — ou du moins de ses convictions affichées — Pasteur ne
pouvait pas penser immédiatement que la vie naissait spontanément de
la matière, qu'elle soit minérale ou organique. La vie devait naître
de la vie. Et, parce qu'il était bien connu qu'un bouillon laissé à
l'air donnait naissance à une activité clairement vivante, il fallait
penser que cela impliquait la préexistence de germes vivants qui se
multipliaient au sein du bouillon. Pasteur pouvait par conséquent
affirmer :
" J'ai la prétention de démontrer avec rigueur que dans toutes les
expériences où l'on a cru reconnaître l'existence de générations
spontanées, chez les êtres les plus inférieurs, où le débat se
trouve aujourd'hui relégué, l'observateur a été victime d'illusions
ou de causes d'erreur qu'il n'a pas aperçues ou qu'il n'a pas su
éviter." (*)
Et comme la vie ne résiste pas aux températures élevées, il devenait facile de détruire tous les germes qui envahissent le bouillon, et, au moyen d'un appareillage approprié, d'empêcher la réinfection du bouillon par des germes vivants. Si cette hypothèse tenait, un bouillon stérilisé par la chaleur resterait inchangé au cours du temps, et ne permettrait pas l'apparition spontanée de la vie. Au contraire, permettre à l'air (où Pasteur suppose que des germes sont présents) d'avoir accès au bouillon suffirait à déclencher le processus bien connu d'apparition d'organismes vivants dans les bouillons ou autres jus, laissés à l'air libre.
La stérilisation de récipients fermés était déjà connue et développée depuis peu (procédé Appert), mais certains objectaient que le principe vital pouvait être de nature très subtile, et n'être en mesure de pénétrer que dans des récipients ouverts. Cela pouvait être testé en utilisant des récipients ouverts à ce principe immatériel, mais où l'ouverture ne permettrait pas à un germe matériel de parvenir aisément au bouillon lui-même. C'est ce que Pasteur réalisa (impliquant qu'il pensait que les germes avaient une masse suffisante pour sédimenter dans le bouillon, et ne pourraient remonter contre la gravité) en construisant ses fameux récipients à col de cygne, ouverts à l'air, mais toujours stériles un siècle après. Il donnait le départ, par ces travaux, à une recherche toujours active sur l'origine de la vie, et au procédé industriel qui porte désormais son nom, la pasteurisation. Mais avons nous beaucoup progressé aujourd'hui ? Quelles sont donc les questions qui restent en suspens ?
Un commencement impossible, la soupe originelle
Malgré la démonstration faite par Pasteur qu'aucune vie ne peut naître spontanément dans un bouillon, c'est le modèle de la "soupe prébiotique" qui a aujourd'hui encore la faveur du public. Mais il y a aussi, hélas, toujours moyen d'échapper à cette question d'origine. Il suffit de la reculer dans des endroits inconnus ou inaccessibles, et d'imaginer que la vie s'est propagée dans l'espace. C'est l'hypothèse de la panspermie. Comme cette hypothèse conduit à une ignorance quasi-absolue, de principe (puisqu'il suffit de reculer, à chaque fois qu'on le souhaite le lieu de l'origine), nous ne la mentionnons qu'en passant.Mais qu'en est-il d'un bouillon originel ? L'idée sous-jacente est qu'une abondante chimie spontanée du carbone a lieu sur Terre et dans l'espace interstellaire. Au fur et à mesure que nous observons le cosmos (et les météorites qui parviennent à terre), il se confirme bien que la chimie du carbone est ubiquiste dans l'univers. Cela aurait permis l'accumulation d'une concentration importante dans les océans d'une grande variété de dérivés organiques, constituant le milieu au sein duquel serait née la vie. Est-il imaginable d'évaluer expérimentalement cette hypothèse, sachant que le déroulement de la vie au cours des ères géologiques a nécessairement totalement brouillé les cartes, en utilisant la matière organique disponible ?
Pour rendre compte de l'évolution d'un riche bouillon originel, Horowitz, en 1945, a proposé l'hypothèse suivante : si la vie utilisait une molécule originelle compliquée, E, elle l'a rapidement épuisée dans ses constructions. Il a donc été nécessaire d'élaborer E à partir d'un précurseur D, à l'aide d'une première catalyse (enzyme a). A son tour D s'est épuisée et il a fallu l'obtenir à partir d'une molécule plus simple C, au moyen d'une enzyme b, et ainsi de suite jusqu'à la molécule la plus simple A. C'est ce qui expliquerait qu'aujourd'hui il existe des organismes vivants qui utilisent les éléments minéraux les plus simples pour se multiplier. Si cette séquence d'événements est exacte on peut donc prédire que b dérive de a, c de b etc., et donc que la phylogénie des enzymes d'un chemin métabolique actuel (procédant du simple vers le compliqué) devrait donner une filiation en sens opposé au sens de la complication des produits du chemin. A l'époque cette hypothèse ne pouvait pas être testée expérimentalement. Elle a encore aujourd'hui la faveur de certains scientifiques (du moins de ceux qui n'ont pas sérieusement réfléchi à la question).
Or, en 1957, Granick proposa une hypothèse diamétralement opposée, où l'évolution se serait faite du simple vers le compliqué, les enzymes évoluant par spécification progressive de leur activité, à partir d'un fonctionnement initial très flou. Les premières activités enzymatiques, dans ce cas, seraient très peu spécifiques et s'accommoderaient de substrats variés. C'est au cours de l'évolution qu'elles deviendraient de plus en plus précises et spécifiques, pour atteindre ce qu'on connaît aujourd'hui.
Ce n'est qu'au début des années 1980, avec les travaux de B.P. Nichols et de ses collègues à l'Université de Chicago, qu'un début de réponse expérimentale a pu être trouvé : ces auteurs, grâce au séquençage des gènes ont pu montrer que l'hypothèse de Granick était la plus vraisemblable. Il appelèrent ce phénomène d'un joli nom, l'"évolution acquisitive". Depuis, de nombreux travaux ont conforté cette hypothèse, mettant peut-être définitivement en défaut celle d'Horowitz. La dernière observation en date, et mettant en jeu des molécules compliquées vient de notre laboratoire, où nous avons montré que des métabolites aussi différents que l'aspartate et le glutamate (précurseurs des protéines), l'uridylate (précurseur des acides nucléiques), et le carbamate (molécule très simple comportant les principaux ingrédients des molécules de la vie), pouvaient être phosphorylés par une même famille enzymatique, l'ancêtre potentiel correspondant à la molécule la plus simple, le carbamate. De même, nous avions montré quelques années auparavant, que le métabolisme de deux autres acides aminés, le tryptophane et la cystéine, proviennent d'un même ancêtre. Cela est remarquable, nous y reviendrons, si l'on se souvient que ces deux acides aminés chimiquement très différents, sont codés par des codons très voisins (même case du code génétique).
Mais de toutes façons il manque à la soupe originelle des molécules essentielles. On n'y trouve pas plusieurs acides aminés (les acides aminés basiques, ceux qui interagissent avec les acides nucléiques), les coenzymes (molécules relativement compliquées, responsables de l'activité catalytique de bien des enzymes), les lipides nécessaires à la confection des membranes, et surtout, les nucléotides (précurseurs nécessaires des acides nucléiques). Cette dernière absence est particulièrement remarquable, en particulier parce qu'il s'agit de molécules très instables, et pour lesquelles il est donc nécessaire de découvrir un processus de synthèse constant, pendant une longue période (ce qui n'est le cas pour l'instant d'aucun processus découvert). Cela reste une des plus grandes énigmes de l'origine de la vie. Nous y reviendrons.
Enfin, la soupe originelle est empoisonnée, car alors qu'il y manque des ingrédients essentiels, elle est remplie de molécules voisines des molécules qu'on trouve aujourd'hui dans les organismes vivants. Or c'est le fait d'être voisin d'un composé normal qui fait qu'une molécule est un poison. Elle prend sa place, s'y substitue, et conduit soit à l'arrêt du métabolisme, soit à sa déviation erronée…
Faut-il une origine à la dissymétrie ?
Pasteur l'a résumé ainsi "la dissymétrie, c'est la vie". Il est donc facile de comprendre que bien des penseurs (du moins ceux qui regardent la surface des choses…) aient imaginé qu'il faut, pour comprendre la vie, expliquer l'origine de la dissymétrie. Et l'on voit fleurir, presque tous les jours, comme c'est le cas pour l'invention du mouvement perpétuel ou la quadrature du cercle, des théories qui expliquent la dissymétrie des acides aminés (dans les protéines ils sont tous d'un seul type), et, partant, de toutes les molécules des organismes vivants. Toutes sortes de conséquences des propriétés de la physique (du monde quantique en particulier) sont censées expliquer cette dissymétrie, à partir d'une dissymétrie essentielle de la physique. Mais c'est justement là que le bât blesse. Pourquoi ne pas reporter à la biologie le raisonnement qui a conduit, en physique, à expliquer la dissymétrie ?Ce raisonnement est simple. La symétrie d'un système physique n'est pas nécessairement un état stable du système. C'est que, contrairement à l'intuition, la symétrie peut être beaucoup plus rare, beaucoup plus coûteuse en organisation ou en énergie, que la dissymétrie. Imaginons une aiguille cylindrique homogène, et imaginons qu'une force s'exerce dans le sens de sa longueur, pour la comprimer. L'expérience montre que, plutôt que se comprimer, l'aiguille va se tordre, dès que la force exercée est un peu forte, perdant ainsi sa symétrie cylindrique. Et l'explication est simple : il faudrait un contrôle infiniment précis du sens de la force exercée pour que l'aiguille garde en se comprimant sa symétrie. La moindre perturbation, même très petite, va entraîner la flexion de l'aiguille. Et cette situation, une fois formée, sera stable. De la même manière, si l'on considère la conduite sur les chemins et les routes, il faut, pour éviter les accidents, conduire ou bien à droite, ou bien à gauche. Et le choix, une fois fait, reste fixé. A moins d'une action dictatoriale, ou concertée : c'est ainsi que Napoléon fit changer le sens de la conduite (on conduisait à gauche en Europe, sauf en Angleterre, bien sûr). Et cette analogie va plus loin : si l'on conduisait de préférence à gauche, c'est que les hommes sont droitiers (cela permettait de se défendre à l'épée contre d'éventuels agresseurs arrivant en face). C'est donc d'une dissymétrie du vivant que provenait le choix du sens de la conduite.
Ainsi il n'est pas impossible qu'une cause physique soit à l'origine de la dissymétrie observée dans le vivant. Mais rien ne le dit : elle est nécessairement très faible, et n'importe quel événement contingent pourrait avoir joué le jeu qu'a joué Napoléon dans la conduite. Car une fois une dissymétrie locale formée, elle s'entretient elle-même. Aussi est-il raisonnable de considérer que l'origine de la dissymétrie du vivant provient de ce qu'elle est simplement plus stable que la symétrie correspondante, mais que savoir pourquoi elle est à gauche plutôt qu'à droite restera sans doute toujours une énigme — de peu d'importance, il est vrai, car, que la forme soit droite ou gauche ne change rien à ce qui fait la vie !
Nous avons mentionné toute une série de molécules, acides aminés, nucléotides, enzymes, dont le lecteur qui se souvient du lycée a une vague idée. Elles ont toutes trait à la vie, mais comment y sont-elles liées ? Il ne peut pas être question ici de résumer le célèbre petit livre de Schrödinger, ni de tenter de définir dans le détail les lois de la vie. Mais je souhaite mettre l'accent sur quatre processus qui sont intimement associés chez tous les organismes vivants et qu'il conviendra donc d'expliquer dans tout modèle de l'origine de la vie. Ce sont : le métabolisme, la compartimentation, la mémoire, et ce qu'on peut appeler la manipulation. Mémoire et manipulation sont reliées par un processus de codage, lui aussi essentiel à la définition de la vie. Concrètement le support physique de la mémoire héréditaire relève de la chimie des acides nucléiques, et plus précisément des Acides DésoxyriboNucléiques (ADN). Il s'agit là de molécules géantes, des macromolécules, et qui sont constituées d'un répertoire de quatre motifs apparentés, les nucléotides, s'enchaînant comme le font les lettres d'un texte alphabétique. Physiquement cet enchaînement est une structure filamenteuse, repliée dans l'espace sous la forme d'une hélice. Plus précisément la molécule d'ADN est formée de deux hélices entrelacées dont la séquence des bases est complémentaire, comme on le verra plus loin.L'information portée par l'ADN dirige la synthèse des protéines, molécules d'une classe chimique différente. Celles-ci sont douées de propriétés manipulatrices. Elles permettent les interconversions du métabolisme et les constructions nécessaires à la compartimentation. Ainsi, les réactions du métabolisme sont catalysées par les protéines. Ces dernières forment aussi un enchaînement linéaire, semblable à celui d'un texte alphabétique, mais de vingt motifs apparentés et non plus de quatre, les acides aminé. Le fil correspondant se replie dans l'espace de façon beaucoup plus compliquée que ne le fait l'ADN, et forme une architecture spécifique de chaque séquence d'acides aminés, et responsable de la fonction de chaque protéine. Dans ces processus deux échelles spatiales, celles des petites molécules (quelques atomes) et celles des macromolécules (quelques milliers, millions ou même milliards d'atomes) sont entrelacées. La vie se déroule principalement à l'échelle intermédiaire, l'échelle mésoscopique.
Ces quatre processus sont de nature si différente qu'il a parfois paru impossible de les associer. Cela a même conduit un physicien comme F. Dyson a proposer que la vie est née deux fois ! C'est ce qui explique pourquoi la plupart des biologistes moléculaires ont tout simplement omis de prendre en compte le métabolisme et la compartimentation dans leurs hypothèses sur l'origine de la vie, pour ne considérer que les protéines et les acides nucléiques. Le même oubli s'est produit chez ceux qui ont poursuivi une réflexion conceptuelle sur la vie, à partir des machines de Turing par exemple, lorsqu'ils ont recherché des programmes capables de produire des images qui leur soient semblables, et qu'ils ont parlé de vie artificielle.
Tout à l'opposé — et cela est le signe d'une surprenante carence intellectuelle de la pensée scientifique — on trouve des chercheurs qui réagissent de façon enthousiaste à la moindre découverte d'une nouvelle molécule organique dans le cosmos, aussi banale qu'un acide aminé par exemple, comme si cela nous apportait une nouvelle explication de l'origine de la vie ! A une autre échelle, la découverte récente des ribozymes, ces acides nucléiques (des ARN, de structure chimique voisine, mais distincte, et beaucoup plus réactive des ADN) doués d'activité catalytique, a été interprétée comme permettant de résoudre l'apparent paradoxe de l'œuf et la poule : qui est le premier, acide nucléique ou protéine ? Mais même si cette question est en effet précisée grâce à l'existence des ARN catalytiques, elle ne fait que poser avec plus d'acuité encore, une question toujours évacuée par ceux qui parlent de l'origine de la vie, celle de l'existence des nucléotides. Il reste par ailleurs une question cruciale, celle de l'origine des membranes, nécessaires à la compartimentation.
Pour comprendre l'origine de la vie il est essentiel de distinguer la reproduction de la machinerie cellulaire, avec ses compartiments (ce que les biologistes "synthétiques" nomment le "chassis") et son métabolisme, et la réplication du programme génétique. Dans son petit livre Origins of Life, Dyson a montré de façon convaincante que dans tout scénario des origines la reproduction doit avoir précédé la réplication. En Biologie Synthétique, les efforts de la plupart des chercheurs sont intéressés par le programme, pas par la machine. Pourtant quelques uns, comme Doron Lancet ou Pier Luigi Luisi, visent à comprendre la phase de reproduction, en construisant des modèles mathématiques ou expérimentaux de ce qui aurait pu se produire dans le passé.
Comment trouver un commencement ? Granick avait proposé d'analyser le métabolisme actuel, pour imaginer le métabolisme originel. Le postulat sous-jacent est ici que c'est à partir de l'origine du métabolisme qu'on pourra expliquer l'apparition et l'organisation de tous les processus du vivant. En bref, à la différence de Cairns-Smith et de beaucoup d'autres il pensait que nous étions en face d'archives, et non de palimpsestes.
La chimie à la surface des pierres
Que faire si l'on écarte dès le départ le modèle de la soupe prébiotique ? De nombreux savants ont proposé une hypothèse alternative. Ils ont insisté sur le fait que pour expliquer l'origine de la vie, nous devrions plutôt considérer un monde minéral, et plus particulièrement la surface de solides, comme lieu des premières réactions qui ont conduit à la vie, plutôt qu'une solution de molécules organiques. Celle-ci nous l'avons vu, n'explique l'apparition ni des acides aminés basiques, ni des coenzymes, ni des lipides nécessaires à la fabrication des membranes, ni des nucléotides…Parmi ces savants on peut retenir quatre noms : Desmond Bernal d'abord, à Londres, qui fit remarquer l'importance des argiles dans la catalyse minérale sur la matière organique ; Samuel Granick, à New York, déjà nommé, qui considéra que l'origine de la photosynthèse devait se trouver sur des surfaces solides, et qui suggéra l'importance de dérivés d'un atome particulier, le soufre, dans les réactions intermédiaires où se transfèrent des électrons (les réactions d'oxydoréduction) ; Graham Cairns-Smith, à Glasgow, qui établit clairement qu'une soupe prébiotique serait littéralement empoisonnée par sa propre richesse en molécules diverses et chimiquement voisines les unes des autres ; et finalement, récemment, Günter Wächtershäuser, au bureau des brevets de Munich, qui insista sur le fait qu'un métabolisme se déroulant à la surface de particules solides devrait être considéré comme la seule possibilité raisonnable pour l'apparition de la vie telle que nous la connaissons. Ce dernier insista aussi sur le rôle du soufre comme intermédiaire principal des réactions d'oxydo-réduction, et comme substrat de nombreuses réactions.
Variations sur un thème : la transformation homéotopique
Brièvement on peut résumer le modèle de ceux qui proposent une origine métabolique, minérale, et solide à la vie, de la manière suivante. Des surfaces minérales convenables, porteuses d'un excès de charges électriques vont garder à leur contact les molécules chargées en sens opposé. Ces molécules sont donc soumises à la fois à un processus de sélection (par la charge électrique) et de concentration locale (elle sont très concentrées à la surface des solides, par le même champ électrique qui les sélectionne). De plus, par simple effet entropique (bien entendu, comme le veut le bon sens, puisqu'on cherche des hypothèses plausibles, sinon probables, en allant dans la direction du deuxième principe de la thermodynamique, et non en sens opposé, comme malheureusement on l'entend trop souvent !), la polymérisation de ces molécules sera favorisée à la surface (en particulier si cette polymérisation élimine une molécule d'eau, qui peut repartir sans contrainte dans le milieu), au contraire de ce qui se passerait en solution.Une justification de cette hypothèse se trouve dans la structure même du métabolisme aujourd'hui, qui met en jeu des molécules riches en phosphates et en carboxylates, donc électriquement très chargées, sans que cela ait une signification chimique pour l'activité qui est la leur dans les processus vivants où elles sont impliquées. De même la plupart des coenzymes, essentiels à la catalyse, sont des molécules ainsi chargées, mais où la charge n'intervient nullement dans leurs propriétés catalytiques (c'est en particulier le cas du pyridoxal phosphate, essentiel à de nombreuses catalyses, où c'est la fonction aldéhyde qui est catalytique, et où le groupe phosphate ne joue aucun rôle).
Pour aller plus avant il est nécessaire de préciser ces hypothèses. Elles ne sont d'ailleurs aujourd'hui toujours que des hypothèses, car la succession exacte des réactions chimiques qui leur correspondent n'a pas été établie en laboratoire. Et le plus rationnel — ce qui fait le moins appel à la magie de l'inconnaissable — est de n'invoquer que des principes simples dans l'évolution primitive. Un principe de ce genre est celui de la variation sur un thème, c'est à dire l'idée que, si une réaction a pu avoir lieu, les réactions semblables auront aussi de bonnes chances de se dérouler. On peut alors prédire l'existence de familles de modifications ordonnées d'un même substrat de base présent fréquemment à la surface du solide, par divers radicaux chimiques. C'est ce que nous avons appelé la transformation homéotopique pour indiquer la conservation d'un thème.
Si cela existe il est important de chercher à dresser le catalogue des réactions possibles, afin de tenter de les réaliser en laboratoire. L'analyse informatisée des gènes (qu'on peut appeler en raccourci analyse in silico, par opposition à l'expérimentation in vivo, ou in vitro, en tube à essais), très nombreux désormais grâce aux programmes de séquençage de génomes entiers, justifie de plus en plus fréquemment l'hypothèse d'une variation sur un thème (homéotopie). On trouve ainsi par exemple que les acides aminés tryptophane ou cystéine pourraient être le résultat d'une transformation homéotopique de la sérine activée. Et cette observation permet d'aller un peu plus loin, en remarquant qu'une modification de ce type existe encore clairement aujourd'hui, pour la synthèse d'un 21e acide aminé, généralement oublié lorsqu'on décrit les protéines, la sélénocystéine. Cet acide aminé rare est présent dans quelques protéines chez des organismes très éloignés les uns des autres du point de vue de l'évolution. C'est une modification par le séléniure d'hydrogène de la sérine portée par un ARN de transfert, acide nucléique servant au décodage du message génétique (une suite de trois nucléotides, ou codon, choisis parmi quatre — notés A, U, G, C — sur le fil de l'acide nucléique, est décodée par l'ARN de transfert qui la reconnaît et place l'acide aminé correspondant dans le fil de la protéine en construction) lorsqu'il est traduit en protéine, qui produit l'acide aminé en question.
Cette observation nous donne alors un fil d'Ariane : si les réactions se déroulaient initialement sur des particules solides, il a fallu un jour les remplacer, puisqu'on ne semble plus les trouver aujourd'hui au sein des cellules. Et l'on est en droit de penser que les ARN de transfert ont été ainsi des substituts des surfaces. Il serait alors aisé de comprendre pourquoi tryptophane (UGG), cystéine (UGC, UGU) et sélénocystéine (UGA) ont des codons voisins : ils correspondent à une même famille de transformations homéotopiques. Mais s'il en est ainsi, il devient intéressant de rechercher les fonctions où apparaissent aujourd'hui des ARN de transfert, en dehors de leur rôle actuel dans la traduction des messagers. Et, de fait, de très nombreux exemples existent de ces fonctions (par exemple pour la synthèse de l'hème, qui donne sa couleur rouge à l'hémoglobine du sang, ou de la chlorophylle, responsable de très nombreux transferts électroniques et de la photosynthèse). Mais parmi ces fonctions il est une transformation homéotopique qui est remarquable, c'est celle qui permet la synthèse de peptides ! En effet des ARN de transfert se trouvent impliqués dans des réactions où ils sont le support de synthèses peptidiques, et cela hors de leur fonction actuelle dans des synthèses de ce type (c'est à dire sans intervention de la machinerie de traduction).
Il devient alors légitime de penser que la synthèse de peptides, en dehors de l'usage du code génétique, a précédé ce qui est aujourd'hui la traduction des ARN messagers, et que c'est sans doute là leur origine. Les messagers ont été d'abord des ARN cofacteurs qui augmentaient la précision de la formation de peptides particuliers, d'où la naissance d'un mécanisme de codage, associant certains acides aminés, à certaines séquences d'ARN. Le code génétique serait donc une conséquence accidentelle de la catalyse par homéotopie de la liaison peptidique.
Il doit paraître clair, à cette description, que nous sommes loin de connaître le détail de l'enchaînement qui a permis la synthèse des protéines. En effet nous avons mis à la suite les unes des autres une série d'hypothèses plausibles, au moins sur le papier, et justifiées, au moins partiellement, par ce qu'on connaît de la vie aujourd'hui. Mais l'enchaînement réel est loin d'être connu. Résumons ce qu'il faudrait identifier. D'abord la nature du solide à la surface duquel se fait le métabolisme primitif. Une argile comme le pense Cairns-Smith ? Mais laquelle ? De la pyrite de fer comme le voudrait Wächtershäuser ? Ensuite on imagine un métabolisme de surface, lequel ? Sans doute de petites molécules porteuses d'un groupe phosphate comme celles qui possèdent trois atomes de carbone, et sont au cœur du métabolisme actuel. Mais il faudrait choisir lesquelles, et dans quelles interconversions. Et ensuite il faut imaginer comment se sont faits les lipides et les membranes, ainsi que les coenzymes. Par ailleurs il est essentiel, avant d'invoquer les ARN comme substituts des surfaces, de savoir comment sont faits leurs précurseurs, les nucléotides. C'est, à notre avis, la question la plus importante à résoudre si l'on veut sérieusement se pencher sur la façon dont est née la vie.
Des peptides qui synthétisent des peptides
Il est en effet bien central — et cela est vrai de toute hypothèse sur l'origine de la vie — de comprendre l'existence des ARN (leur multiplication est devenue plausible, mais non démontrée dans un détail suffisant, depuis qu'on sait qu'il existe des ribozymes, capables à la fois de catalyse et de reproduction). Comme la polymérisation est plausible sur des surfaces, nous l'avons vu, le problème central reste celui de l'existence des nucléotides. C'est un problème très difficile, et généralement gravement sous-estimé. Aucune solution satisfaisante n'existe pour l'instant, en raison à la fois de la très grande complication de ces molécules, et de leur grande instabilité (qui fait qu'il n'est pas raisonnable de penser que ce sont des molécules rares qui se sont peu à peu accumulées).Or, leur métabolisme actuel, très différent de ce que font les chimistes organiciens lorsqu'ils synthétisent ces molécules au laboratoire, met en jeu des acides aminés comme précurseurs. On est donc naturellement conduit à privilégier des hypothèses concernant les acides aminés, et les peptides. Existe-t-il des réactions actuelles qui permettent la synthèse de peptides hors de la traduction ? Depuis quelques années on sait que c'est le cas, autour d'un coenzyme (phosphorylé et utilisant le soufre pour son action catalytique, comme on s'y attend si l'on a suivi ce qui précède) le 4-phosphopantéthéine. Ce coenzyme transporte des acides aminés chimiquement activés, sur une suite d'atomes de soufre, au travers d'une séquence de transferts de liaisons thioesters. Et cela se justifie bien : le chimiste Christian De Duve a donné de nombreux arguments pour proposer que la liaison thioester ait été primordiale, dans la chimie de l'origine de la vie.
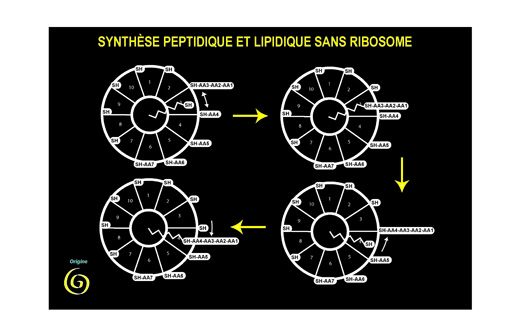
Les réactions qui mettent en jeu ce coenzyme permettent la synthèse de peptides de structures variées (en général des antibiotiques). Plusieurs aspects de la réaction sont archaïques, en particulier le fait que les acides aminés sont soit droits ou gauches, ou le fait que la liaison mise en jeu soit une liaison sur un atome de soufre. Il devient ainsi remarquable qu'un (iso) peptide serve à la synthèse de (iso) peptides. Bien sûr l'hypothèse serait fortement confortée si l'on trouvait dans un organisme quelconque que le 4-phosphopantéthéine est synthétisé à partir d'une enzyme qui l'utilise comme cofacteur. Cela n'est pas encore le cas.
Mais il y a une observation (faite par Fritz Lipmann à propos de l'organisation des structures enzymatiques correspondantes, bien avant que les séquences protéiques soient connues) qui rend cette hypothèse encore plus remarquable : l'analyse de la séquence des gènes permettant la synthèse des acides gras, lipides constituants majeurs des membranes, a montré que ce sont des enzymes de la même famille (et avec le même coenzyme) qui permettent leur synthèse. Nous aurions là, une première explication de la genèse des membranes (essentielles à la compartimentation), comme sous-produit accidentel de la synthèse des peptides…
La fixation de l'azote et la synthèse des nucléotides
On sait aujourd'hui que l'atmosphère terrestre primitive n'était pas très réductrice, et il est donc peu probable qu'il y ait eu une forte concentration de NH3 il y a 3,8 milliards d'années. L'atmosphère était pour l'essentiel riche en CO2 et N2, et peut être en dérivés plus oxydés de l'azote. Si donc nous faisons l'hypothèse de l'existence d'acides aminés assez abondants il faut imaginer un mécanisme efficace de fixation de l'azote au sein de molécules à squelette carboné. C'est là encore une question essentielle, non résolue de façon satisfaisante, occultée par la plupart des chercheurs qui s'interrogent à propos de l'origine de la vie.En suivant l'approche de Granick il nous faut pour progresser, considérer la façon dont se fixe l'azote aujourd'hui. En général cela requiert la présence de protéines contenant un noyau fer-soufre (comme dans les ferrédoxines, nécessaires à de très nombreux transferts électroniques) pour assurer les transferts d'électrons intermédiaires, et du molybdène pour la fixation de l'azote de l'air. Dans les cas d'oxydations plus complexes, un cofacteur comprenant une molybdoptérine, et du molybdène pour les transferts d'électrons est retrouvé de façon très fréquente. Le détail général de ces transferts électroniques n'est pas encore bien connu, et il existe une assez grande variété de mécanismes d'assimilation de l'azote suivant les organismes considérés.
Les ferrédoxines sont de très petites protéines formées d'un nombre limité d'acides aminés différents, comme on l'attendrait de protéines ancestrales, et elles contiennent le noyau fer-soufre dont l'existence est postulée par Wächtershäuser comme fondamental à l'origine (pour lui la pyrite de fer est un des minéraux cruciaux pour le métabolisme primitif). Le molybdène est un élément très rare de la croûte terrestre, mais il n'est pas clair que cela a toujours été le cas. Par ailleurs d'autres métaux auraient pu jouer le même rôle dans une atmosphère sans oxygène (c'est le cas du tungstène encore aujourd'hui).
Mais la molybdoptérine est aussi un coenzyme contenant du soufre (ce qui lui permet de se fixer au noyau fer-soufre, ou à la pyrite), synthétisé à partir d'un précurseur nucléotide, le GTP, par cyclisation avec perte d'un groupe à un carbone, le résidu formyl. Cette étape est catalysée par la GTP cyclohydrolase, qui forme la ptéridine triphosphate à partir du GTP, et libère un résidu formyl (par ailleurs transporté par des coenzymes dont le cœur est une ptéridine). Ne pourrait-on pas alors envisager la réaction inverse, qui serait un moyen direct de former les nucléotides ?
PteridineTP + HCOOH -----> GTP + H2O
Nous aurions alors à faire l'hypothèse qu'un processus autocatalytique assurerait la synthèse de ptéridine-phosphate, nécessaires à la fixation de l'azote, et que la synthèse des nucléotides ne serait qu'un sous-produit accidentel du défaut de précision de cette synthèse. Ainsi le GTP ne serait qu'un sous-produit de la fixation de l'azote.
Cette hypothèse bien hardie demande que l'on s'interroge sur la synthèse des ptéridines à partir d'(iso) peptides. Mais l'exploration du métabolisme microbien qui se développe à grande échelle autour des programmes de séquençage des génomes entiers devrait nous permettre bientôt de voir si la direction ainsi esquissée est intéressante.
Conclusion provisoire
L'abandon de l'hypothèse d'un bouillon riche en molécules organiques à l'origine de la vie, et son remplacement par l'hypothèse d'un métabolisme de surface se justifie de mieux en mieux au travers de ce qu'on commence à entrevoir de la nature des génomes. La vie forme un tout, et il n'est donc pas possible d'oublier métabolisme ou compartimentation comme en faisant partie de façon intime. C'est d'ailleurs l'idée d'une chimie précurseur du métabolisme actuel qui fournit les fils d'Ariane qui nous permettent de retrouver un scénario d'origine. Au fur et à mesure que sont connus de nouveaux gènes il apparaît qu'au contraire de ce qu'on pouvait craindre, nous sommes sans doute beaucoup plus près de l'origine — malgré les trois milliards et demi d'années d'évolution — que nous ne le pensions. Mais cela montre en même temps de façon particulièrement visible combien nous sommes loin de comprendre l'origine de la vie, alors qu'il existe des dizaines de scénarios différents, proposés de façon très fragmentée, et sans jamais être intégrés dans une vision globale par de nombreux chercheurs à travers le monde. Toute la question est de savoir si l'origine a été effacée par la vie actuelle (c'est ce que pense G. Cairns-Smith, qui parle alors de relève génétique, des processus minéraux ancestraux ayant été relevés — comme on relève la garde d'un fortin — par les processus actuels, ou S. Benner, qui parle de palimpseste) ou si elle est au contraire encore bien visible dans la vie actuelle (c'est ce que pensait S. Granick, ou G. Wächtershäuser aujourd'hui et qui semble encore difficile à comprendre par beaucoup). Il va de soi que, dans le premier cas, la plupart des recherches sont, par principe, vouées à l'échec, et qu'il est donc plus intéressant, même si cela doit, au cours du temps se révéler inefficace, de favoriser la deuxième façon de voir, et de tenter de comprendre, par la vie actuelle, sa naissance.Parmi les nombreuses conséquences conceptuelles des directions ainsi indiquées, enfin, on peut remarquer un nouveau regard sur la chimie, où les surfaces jouent un rôle de premier plan, mais aussi où l'idée de sélection est centrale. La chimie sélective pourrait être non seulement pour les laboratoires, mais pour l'industrie, une chimie du futur. Et l'hypothèse de Granick, qui veut que la sélectivité ait été progressive, devrait être pour nous un modèle si nous voulons imaginer une chimie du futur qui soit biomimétique, sans pour autant qu'elle ait à expliquer l'origine de la vie.
Notes
(*) Louis Pasteur
"Sur les corpuscules organisés qui existent dans l'atmosphère. Examen
de la doctrine des générations spontanées" Leçon professée à la
Société chimique de Paris, le 19 mai 1861.
in : Leçons de chimie et de physique professées en 1861 (à la Société
chimique de Paris), Paris, 1862, Hachette et Cie, p. 219-254 (retour
au texte)
Bibliographie sommaire
Bernal JD (1951) The physical basis of life. Routledge and Kegan
Paul, Londres.
Brack A & Raulin F (1991) L'évolution chimique et l'origine de la
vie. Masson, Paris.
Cairns-Smith, G. (1982) Genetic takeover and the mineral origin of
life. Cambridge University Press, Cambridge.
Crick F (1982) La vie vient de l'espace. Hachette, Paris.
Danchin A (1990) Une aurore de pierres. Aux origines de la vie. Le
Seuil, Paris.
De Duve C (1991) Blueprint for a cell. Neil Patterson, Burlington,
NC/Portland Press, Londres.
Dyson FJ (1985) Origins of life. Cambridge University Press,
Cambridge.
Granick S (1957) Speculations on the origin and evolution of
photosynthesis. Annals of the New York Academy of Sciences 69,
292-308.
Schrödinger E (1944) What is life? Traduction française : Qu'est ce
que la vie ? (1986) Bourgois, Paris.
Wächtershäuser G (1988) Before enzymes and templates : theory of
surface metabolism. Microbiological Reviews 52, 452-480.