Pour
aller directement au contenu principal : Pressez
Entrez
Philosophie Zoologique
Jean-Baptiste de Lamarck
L'aurore des pierres
Conférence du Centre International de Synthèse
Collège de France, 7 février 1990
Jacques Roger (1920-1990) était un philosophe et historien des sciences qui veillait avec rigueur à ne pas succomber à la mode ou à la pensée molle, cette pensée superficielle qui fait le cœur désormais des quotidiens qui se prennent pour vecteurs de l'intelligence et la stérilisent. Il avait organisé au Collège de France une série de conférences dans la lignée de ce qu'avait proposé le philosophe Henri Berr (1863 – 1954), le créateur de la Revue de Synthèse, fondée sur un renouvellement de l'Histoire, de l'histoire des idées en particulier. Son actualité pour la biologie d'aujourd'hui, qui se fait biologie de synthèse (la moderne "biologie synthétique", qui reprend le mot de Stéphane Leduc), est réjouissante.
Le texte de cette conférence est la transcription d'une expression orale, avec les redites et les approximations spécifiques à ce mode d'expression (qui ne se relit pas). Il devait être publié dans un ouvrage collectif sous la direction de Jacques Roger, qui, frappé par la mort de façon soudaine à peine deux mois après cette conférence, n'a pu le mener à son terme. Ce texte reflète l'état de ma pensée au début de l'année 1990 au moment où je proposais le terme in silico, pour exprimer le mode expérimental, complémentaire de l'approche in vivo et de l'expérience in vitro, utilisant les ordinateurs pour explorer les conjectures que fait l'esprit humain pour représenter le réel (1). Je me faisais alors l'avocat des programmes de séquençage des génomes, mais pas à la manière d'une prouesse technique, comme cela était présenté pour le génome humain, mais avec un véritable objectif scientifique, en particulier celui de découvrir au sein des génomes les traces de l'orgine de la vie. Dans ce contexte j'analyse le métabolisme et je tente de faire oublier — nous sommes en 1990 — la très mauvaise image, encore en cours aujourd'hui malgré son absurdité, de la "soupe prébiotique". Cette conférence a été développée dans un livre publié aux Editions du Seuil, Une Aurore de Pierres. Certains aspects ont été développés dans l'Enciclopedia Einaudi, dirigée par Ruggiero Romano (1923-2002), jusqu'à sa disparition.
Mesdames, Messieurs, nous voici à la troisième conférence de cette série. Je suis heureux, ce soir, de pouvoir vous accueillir dans des conditions un peu moins inconfortables, au moins pour la grande majorité d'entre vous, qu'au cours des conférences précédentes, le Collège ayant pu ce soir nous donner une salle plus grande. Je rappelle à ceux qui nous ont déjà posé la question, que nous envisageons de publier le texte de ces conférences et je demande à ceux d'entre vous qui seraient intéressés par cette publication, de mettre leur nom, la fois prochaine ou la suivante, sur le cahier qui est à l'entrée de la salle. Cela ne signifie pas du tout qu'ils s'engagent à acheter la publication, mais cela signifie qu'ils seront tenus au courant de cette publication. Nous avons jusqu'à présent entendu Monsieur Audouze, un astrophysicien, et Monsieur Levy-Leblond, un physicien. Ce soir, c'est un biologiste que nous allons entendre. Il n'est pas nécessaire de présenter Monsieur Antoine Danchin qui est Directeur de Recherche au CNRS et Chef de l'Unité de Régulation de l'Expression Génétique à l'Institut Pasteur. Monsieur Danchin est un généticien moléculaire et c'est un tout autre univers que ceux que nous avons vus dans les conférences précédentes, c'est un tout autre univers que nous allons découvrir ce soir grâce à lui. Je le remercie d'être venu. Je lui donne immédiatement la parole.
Je remercie d'abord Jacques Roger de m'avoir invité devant un public aussi choisi que ceux qui perpétuent ce qu'avait voulu Henri Berr, et je vais tout de suite commencer par un élément qui se relie directement à la tradition de ce qu'avait voulu faire Henri Berr, c'est-à-dire par quelques remarques épistémologiques. Avant de vous présenter le thème de l'origine de la vie, je vais vous montrer un peu la façon dont je vois que se crée la science, et en particulier, la science dans ce domaine précis. En effet, je crois que le réel ne parle pas. C'est là un point assez essentiel qui indique que nous ne pouvons que faire des devinettes sur le réel et, comme disait Xénophane, il y a deux mille cinq cents ans, ce sont devinettes car, même si par hasard, nous tombions sur la vérité, nous ne pourrions pas le savoir.
Et ce que je voudrais vous montrer, c'est comment on peut s'inscrire dans la production de modèles du réel, au sein d'un certain nombre de théories qui permettent d'agir sur le réel, d'essayer de le forcer à réagir, puisque lui-même ne parle pas.
En quelques mots, je vais vous présenter un premier schéma très, très simple dans lequel apparaît l'attitude scientifique. Pour moi, cette attitude, c'est d'abord la production de modèles, et j'entends, production de modèles au sens le plus général, ce peut être par exemple produire une statue d'argile à laquelle on fait subir toutes sortes de bienfaits ou de sévices : c'est bien là, pour moi, une attitude scientifique. Ce qui compte essentiellement, donc, n'est pas tellement — puisqu'on constate qu'il existe un progrès dans la façon de se représenter le réel et d'interagir avec lui — ce n'est pas tellement de produire des modèles, mais de produire des modèles qui ont une certaine adéquation au réel, des modèles qui puissent d'une manière ou d'une autre le faire réagir.
Et nous devons, pour le faire, partir de la situation dans laquelle nous sommes, c'est-à-dire d'une culture et d'une histoire. Nous commençons par une question et des idées reçues. Et l'une des raisons pour lesquelles — vous l'avez remarqué peut-être — j'ai volontairement rompu le fil des titres des conférences de cette année est essentiellement parce que ce que je vais chercher à faire d'abord, avant de vous exposer quelques vues sur l'origine de la vie, c'est d'une certaine manière essayer de semer le doute, semer le doute envers des idées reçues. C'est un point assez important, je crois, qui nous permettra d'aborder des questions difficiles et aussi immodestes que les problèmes posés par l'origine de la vie.
Nous partons donc d'idées reçues, et ces idées reçues sont formalisées, plus ou moins bien, et sont plus ou moins décrites en termes de postulats. Un postulat, c'est une façon de formaliser ou d'écrire ces idées reçues. Maintenant, à partir de ces postulats, on va vouloir construire un modèle, et comme je vous l'ai dit, ce modèle peut être une statue d'argile ou, beaucoup plus souvent chez nous, ce peut être un modèle mathématique, par exemple. On doit donc, dans ce dernier cas, interpréter les postulats en axiomes et définitions ; et, à partir de là, on va construire une théorie cohérente à partir d'une logique (elle-même fondée sur une métalogique, à propos de laquelle il y aurait d'ailleurs beaucoup à discuter, parce que, là encore, on est plongé d'une certaine manière dans la culture et dans l'histoire). Eh bien, le modèle lui-même va se dérouler en produisant des théorèmes ou, le plus souvent, malheureusement, des conjectures de théorèmes (c'est-à-dire qu'on ne sait pas en faire la démonstration). En parallèle, d'ailleurs, on réalise souvent quelque chose qui est assez voisin de la statue de glaise : on fait des simulations du modèle, simulations qui sont sensées le représenter. Puis, lorsqu'on est arrivé à construire ces conjectures de théorèmes ou ces simulations de conjectures, on doit retourner vers le réel et via une interprétation, les transposer en prédictions expérimentales. Et ces prédictions expérimentales sont de deux types, ou bien elles sont existentielles, — on prédit l'existence d'un objet on doit l'exhiber, c'est l'objet qu'on doit découvrir, ou bien elles sont d'un tout autre type et elles sont en principe réfutables, c'est-à-dire qu'on peut montrer qu'elles étaient inadéquates.
![]()
Parmi ces idées reçues, il y en a une qui me paraît profondément non scientifique et qui joue un rôle très important dans nos cultures, bien que depuis fort longtemps, il ait été perçu qu'elle n'était pas scientifique, c'est l'idée d'un rôle essentiel du hasard. Pour moi, le hasard n'est pas une donnée scientifique, mais pas non plus d'ailleurs, une notion grecque. Et j'avais été extrêmement surpris par l'épigraphe du livre de Jacques Monod d'où il tirait son titre "Le hasard et la nécessité". Aussi ai-je eu la curiosité de chercher chez Démocrite la citation en question. Elle n'est pas chez Démocrite. Non seulement elle n'est pas chez Démocrite, mais si l'on étudie, avec un peu de soin, les textes pré-socratiques, et ce n'est pas très difficile puisque finalement, ils ne sont pas très nombreux, on trouve une phrase et une seule qui ressemble à la phrase de Jacques Monod, et qui est "Oὐδὲν χρῆμα μάτην γίνεται ἀλλα πάντα ἐκ λόγου τε καὶ ὑπ᾽ ἀνάγκης", ce qui veut dire qu'aucune chose dans la nature ne vient pas soi-même, mais toutes choses sont le fruit d'un logos et de la nécessité. Le logos n'est certainement pas le hasard, et c'est essentiellement cet aspect que je voudrais discuter ; si nous pouvons espérer remonter aux origines, nous devons chercher des lois, chercher des règles de construction ; et si nous les trouvons, alors nous avons quelques chances de remonter aux origines. C'est essentiellement la leçon que je voudrais donner pour commencer. Et comme je ne voudrais pas rester sur une image négative de la contribution de Jacques Monod, contribution qui a été extrêmement positive en fait, je voudrais continuer par une phrase de lui — la précédente était apocryphe, elle était donc de lui, en réalité, évidemment, mais c'était une interprétation de sa part qu'il avait signée Démocrite, qu'il n'a pas signée Mac Grégor, comme à son habitude — mais il a écrit une tout autre chose dans une pièce inédite, dans laquelle il affirme clairement l'existence de lois : "Asservir la nature ?... Etrange expression. Pour y parvenir, Philokratos, il faut d'abord la respecter, l'écouter, lui obéir. C'est ce que j'essaie de faire, maladroitement. Vois cette toupie, je puis la lancer, non l'asservir ; ce n'est pas à moi qu'elle obéit, mais à une loi (un temps), une loi que j'ignore encore" (Jacques Monod, Le Puits de Syène, Inédit).
Cette fois, c'est son attitude de savant qu'on peut voir ici. C'est l'attitude, je dirais, de tout scientifique et qui consiste à chercher des lois. Et la question qui va se poser à nous, pour la biologie, sera : est-ce qu'il existe des lois biologiques et quelles sont-elles ? Avant d'arriver à cette question des lois biologiques, on pourrait être amené à un grand nombre de questions sur la nature des lois physiques etc, etc, mais je voudrais essentiellement vous dire que la biologie, contrairement à ce qu'on pense, parce que nous sommes des êtres biologiques, est une science particulièrement abstraite et non particulièrement concrète et donc, particulièrement difficile à comprendre et non particulièrement facile à comprendre. C'est là un point qui va être très important ; les lois dont je vais vous parler sont des lois extrêmement abstraites ; et la raison en est la suivante : on a tendance, en général, à rechercher des objets, et à se contenter de la découverte d'objets, de leur identification et l'on a l'impression qu'on a compris quand on a trouvé des objets. Eh bien, dans le cas des êtres vivants, leur particularité est que ce ne sont pas tant les objets qui comptent, que les relations entre ces objets, et même les relations entre ces relations. C'est là quelque chose qui est extrêmement abstrait et donc très difficile à comprendre. En général, un certain nombre des relations auxquelles je vais faire allusion, et que je ne vais pas pouvoir détailler, évidemment, en si peu de temps, sont des lois très abstraites qu'en un sens, on peut avoir l'impression intuitivement de comprendre immédiatement. Mais en fait, on se rend compte, très longtemps après, qu'on n'a pas réellement compris. Cela demande beaucoup de profondeur et beaucoup de réflexion, et nous n'en sommes finalement qu'aux balbutiements. La biologie, disons contemporaine, ce que nous pouvons faire en ce moment, a une cinquantaine d'années tout au plus. C'est très peu et, par conséquent, il y a encore énormément à comprendre, énormément à découvrir ; l'âge d'or n'est pas passé, nous en sommes encore tout-à-fait au début.
Parmi les différentes lois biologiques possibles, dont je vais vous donner une petite liste, je vais en retenir seulement deux. Voici, selon moi, quelques-unes de ce qu'on pourrait considérer comme des lois biologiques. Je vais retenir deux de ces lois parce que je les ai déjà mentionnées à propos de la thématique de l'origine. La première qui, à mon sens, est de loin la plus importante, c'est le codage, à savoir essentiellement la séparation entre deux aspects, un aspect fonctionnel des choses, manipulatoire, et un aspect, disons, mémoire des choses. Le fait qu'il existe une relation qui puisse séparer ces deux aspects et les mettre en correspondance crée une boucle qui a des propriétés tout-à-fait originales, et qui sont les propriétés mêmes du vivant. Cela a été inventé deux fois, une fois, c'est ce qui a créé la vie, et la deuxième fois, c'est ce qui a créé ce que nous sommes en train de faire aujourd'hui, c'est le langage. C'est cette aptitude pour un système vivant à revenir sur lui-même et à créer quelque chose de nouveau, d'émergent à partir de ce propre retour sur soi. Un deuxième aspect, beaucoup plus restreint, est ce qu'on peut appeler la relève génétique (2). C'est là une question qui est importante lorsqu'on se pose la question des origines. La voici : lorsqu'on considère aujourd'hui des êtres vivants, et qu'on essaie de découvrir leur généalogie, on peut remonter à quelques ancêtres. Et l'on peut avoir la tentation de penser que les ancêtres devaient en quelque manière être semblables à leurs prédécesseurs, et qu'on conservera toujours la trace des prédécesseurs. En réalité, il peut s'être produit la chose suivante, qu'il est apparu un jour un type particulier de fonctionnement donnant naissance à un type nouveau, et que ce nouveau type a entièrement effacé, parce qu'il possédait des propriétés originales, ce qui était là avant lui. On peut d'ailleurs s'interroger sur la propriété particulière qu'a l'espèce humaine actuellement sur la Terre de rapidement effacer les archives de sa naissance. C'est une des questions évidemment tout-à-fait essentielles.
Nous sommes donc amenés à une
première question qui mériterait évidemment des heures de réflexion,
et qui est tout simplement : qu'est-ce que la vie ? Je vais retenir un
certain nombre de particularités liées à la vie, les quatre processus
que j'ai marqués ici. D'abord, les deux premiers aspects sont souvent
oubliés. Le métabolisme, point tout-à-fait essentiel mais en
général omis, est l'aptitude à transformer chimiquement son
environnement : un être vivant est un transformateur chimique de son
environnement. Il prend dans son environnement un certain nombre
d'objets, il en recrée d'autres ; c'est le métabolisme. Le deuxième
aspect absolument essentiel est qu'il n'y a pas d'être vivant sans
membrane ou sans peau. Un être vivant est constitué d'un intérieur et
d'un extérieur. Ensuite, apparaissent deux aspects sur lesquels a été
en général mis l'accent (mais beaucoup plus récemment). Ce sont, d'une part, l'aptitude à transmettre "quelque chose" de
génération en génération, ce que j'appelle ici mémoire, et
dont le support chimique est la famille des acides nucléiques, et
ensuite, bien sûr, l'aptitude à manipuler l'environnement à
partir d'une transposition de la mémoire en ces objets particuliers
que sont les protéines, au travers de la catalyse réalisée par les
protéines. Ce qu'on remarque ici, dans cette double classification,
c'est que deux échelles d'espace particulières sont mises en relation
entre elles. Là intervient justement une question de relations, comme
je vous l'ai dit au début, d'une très grande importance : on observe
deux échelles différentes, une échelle de petites molécules, typiques
de ce qu'on trouve dans le métabolisme ou dans les molécules qui
constituent les membranes (encore que les membranes créent une échelle
particulière qui est l'échelle de la cellule, ce qui mériterait de
longs commentaires) et puis, une échelle tout-à-fait originale à la
vie, la constitution des macromolécules. Il s'agit là d'enchaînements
de dizaines de milliers d'atomes, enchaînements qui sont tous
constitués à partir d'un petit nombre d'objets de base. Je dois vous
dire aussi que, quand on regarde de près l'ensemble de ces quatre
processus fondamentaux, on s'aperçoit que le nombre des objets qui
permettent leur mise en place est très restreint. Le métabolisme, par
exemple, dans une cellule entièrement autonome, comprend environ cinq
cents types chimiques différents, pas plus. On peut identifier tous
les métabolites, c'est tout-à-fait accessible à l'expérience et peut
s'écrire sur un tableau qui n'est pas tellement long. On peut aussi
décrire, d'ailleurs, les flèches qui relient ces objets les uns aux
autres. De la même manière, les acides nucléiques sont faits d'un
enchaînement de quatre ou cinq types de molécules voisines
chimiquement, et uniquement de cela. C'est-à-dire que ces éléments de
base s'enchaînent comme des lettres de l'alphabet qu'on peut écrire
sous forme de texte. De même pour les protéines, mais ce sont, cette
fois-ci, vingt (ou vingt et un)(3)
acides aminés qui permettent de fabriquer l'enchaînement des
protéines. Ce sont d'abord ces aspects-là que je vais retenir de la
vie. Et la question qui se pose à nous, puisque c'est la question de
l'origine, est d'imaginer comment s'est créé cet ensemble. De façon
habituelle, et jusqu'à très récemment, la plupart des auteurs ont
essentiellement raisonné sur les deux derniers aspects de la question,
à savoir l'origine des protéines et des acides nucléiques. Mais la
question qui se pose alors est toujours une question du type de l'oeuf
et de la poule : qui est premier, la protéine ou l'acide nucléique,
puisque l'acide nucléique est la mémoire qui permet de faire la
protéine et que la protéine est nécessaire pour faire l'acide
nucléique ? Ce que je vais essayer de vous montrer aujourd'hui, c'est
qu'en fait on peut reformuler la question de façon différente en
écartantces deux éléments de départ et en cherchant l'origine dans le
métabolisme. Et ce que je vais vous proposer, c'est que
l'origine apparaît grâce à un type métabolique particulier qui va créer
cet ensemble.
Ce sont, d'une part, l'aptitude à transmettre "quelque chose" de
génération en génération, ce que j'appelle ici mémoire, et
dont le support chimique est la famille des acides nucléiques, et
ensuite, bien sûr, l'aptitude à manipuler l'environnement à
partir d'une transposition de la mémoire en ces objets particuliers
que sont les protéines, au travers de la catalyse réalisée par les
protéines. Ce qu'on remarque ici, dans cette double classification,
c'est que deux échelles d'espace particulières sont mises en relation
entre elles. Là intervient justement une question de relations, comme
je vous l'ai dit au début, d'une très grande importance : on observe
deux échelles différentes, une échelle de petites molécules, typiques
de ce qu'on trouve dans le métabolisme ou dans les molécules qui
constituent les membranes (encore que les membranes créent une échelle
particulière qui est l'échelle de la cellule, ce qui mériterait de
longs commentaires) et puis, une échelle tout-à-fait originale à la
vie, la constitution des macromolécules. Il s'agit là d'enchaînements
de dizaines de milliers d'atomes, enchaînements qui sont tous
constitués à partir d'un petit nombre d'objets de base. Je dois vous
dire aussi que, quand on regarde de près l'ensemble de ces quatre
processus fondamentaux, on s'aperçoit que le nombre des objets qui
permettent leur mise en place est très restreint. Le métabolisme, par
exemple, dans une cellule entièrement autonome, comprend environ cinq
cents types chimiques différents, pas plus. On peut identifier tous
les métabolites, c'est tout-à-fait accessible à l'expérience et peut
s'écrire sur un tableau qui n'est pas tellement long. On peut aussi
décrire, d'ailleurs, les flèches qui relient ces objets les uns aux
autres. De la même manière, les acides nucléiques sont faits d'un
enchaînement de quatre ou cinq types de molécules voisines
chimiquement, et uniquement de cela. C'est-à-dire que ces éléments de
base s'enchaînent comme des lettres de l'alphabet qu'on peut écrire
sous forme de texte. De même pour les protéines, mais ce sont, cette
fois-ci, vingt (ou vingt et un)(3)
acides aminés qui permettent de fabriquer l'enchaînement des
protéines. Ce sont d'abord ces aspects-là que je vais retenir de la
vie. Et la question qui se pose à nous, puisque c'est la question de
l'origine, est d'imaginer comment s'est créé cet ensemble. De façon
habituelle, et jusqu'à très récemment, la plupart des auteurs ont
essentiellement raisonné sur les deux derniers aspects de la question,
à savoir l'origine des protéines et des acides nucléiques. Mais la
question qui se pose alors est toujours une question du type de l'oeuf
et de la poule : qui est premier, la protéine ou l'acide nucléique,
puisque l'acide nucléique est la mémoire qui permet de faire la
protéine et que la protéine est nécessaire pour faire l'acide
nucléique ? Ce que je vais essayer de vous montrer aujourd'hui, c'est
qu'en fait on peut reformuler la question de façon différente en
écartantces deux éléments de départ et en cherchant l'origine dans le
métabolisme. Et ce que je vais vous proposer, c'est que
l'origine apparaît grâce à un type métabolique particulier qui va créer
cet ensemble.
Alors, puisque je vous ai dit au tout début que nous aurions à revenir sur des idées reçues, nous devrons d'abord en écarter une, extrêmement répandue — est-ce que c'est Darwin qui en est la cause avec sa "petite mare tiède", je ne sais pas — l'idée que, à l'origine, il existe un bouillon de culture, une soupe prébiotique. Cette idée moyen-âgeuse est extrêmement répandue, peut-être parce qu'on voit assez facilement que les bouillons de culture se contaminent et deviennent "vivants" rapidement : mais on sait très bien que Pasteur avait déjà démontré qu'ils ne pouvaient devenir vivants que si la vie était déjà présente. C'est, à cause de cette idée de bouillon prébiotique, que les gens sont allés chercher les molécules du système vivant un peu partout, dans l'espace interstellaire, etc, etc, et les ont découvertes sans d'ailleurs grande difficulté. On a démontré ainsi que la chimie du carbone, la base de la chimie du vivant, est une chimie répandue. Mais ce qui a été rarement remarqué, c'est qu'on ne trouve pas du tout n'importe quoi, un peu partout. Deux phénomènes sont frappants. D'abord, on trouve beaucoup plus de types de molécules que celles qui se trouvent dans le système vivant. Ensuite, il manque un très grand nombre de molécules pourtant tout-à-fait essentielles. Et pour celles-là, on n'a aucune idée de leur mode de synthèse. Ces molécules essentielles sont quelques acides aminés très importants, parce que ce sont les acides aminés qui se lient aux acides nucléiques ; ce sont aussi des bases des acides nucléiques, la moitié des acides nucléiques, pourrait-on dire ; ce sont encore la plupart des co-enzymes, petites molécules qui permettent la catalyse réalisée par les protéines et ce sont enfin les nucléotides permettant la mise en place des enchaînements des acides nucléiques. Enfin, bien sûr, je vous ai mentionné les membranes : on ne sait pas réaliser les lipides. Tout cela veut dire que le bouillon prébiotique est peut-être très riche, mais il y manque des ingrédients essentiels. Et cette richesse même conduit à une réflexion qu'on peut avoir sur l'idée de poison. Qu'est-ce qu'un poison aujourd'hui ? Un poison, c'est une molécule d'un type particulier qui ressemble à une molécule du vivantet prend sa place. Comme elle prend sa place, et qu'elle ne lui est pas identique, elle arrête le fonctionnement du système dans un endroit ou un autre et a donc une action délétère. Or, justement, c'est la situation que l'on trouve dans les expériences qui ont été réalisées avec l'idée d'un bouillon prébiotique. Dans ces expériences où l'on reconstitue une atmosphère primitive, où l'on essaie d'imaginer qu'on avait dans l'atmosphère primitive, des étincelles, des orages, ou des rayons ultra-violets, tout ce qu'on veut comme forme d'énergie, on reconstitue des mélanges du type suivant.
Voici
une expérience typique de Miller. 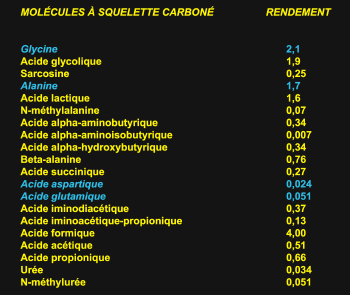 Ce
que montre cette liste, c'est que, dans un mélange prébiotique, avec
une atmosphère imaginaire, on trouve un grand nombre de molécules plus
ou moins orientées. Et parmi ces molécules, quatre seulement sont des
acides aminés qui participent aux protéines aujourd'hui. Ainsi, l'un
des points tout-à-fait clair qui dérive de l'idée de bouillon
prébiotique, c'est qu'en fait on a un bouillon empoisonné, beaucoup
trop riche, mais dans lequel il manque des ingrédients essentiels.
C'est à ce stade qu'il devient intéressant de se demander comment on
pourrait imaginer à la fois un système qui puisse engendrer de la
variété mais surtout, introduire une sélection. C'est un point
essentiel sur lequel j'insiste parce que l'idée de sélection est une
idée très extrêmement répandue dans le monde anglo-saxon, mais
curieusement, très peu répandue en France où nous avons une tradition
assez particulière dans ce domaine. Quelle serait la sélection, le tri
assez efficace, pour ne retenir que certaines classes de
molécules ? Et ce sont ces molécules qui, elles, doivent pouvoir
constituer le premier métabolisme et engendrer un certain nombre des
éléments de base qui permettront la synthèse des êtres vivants. A ce
propos, un Anglais, Graham Cairns-Smith, depuis assez longtemps,
reprenant des idées antérieures, a eu une réflexion extrêmement
intéressante sur le rôle possible des argiles dans l'origine de la
vie. Il a écrit un livre, Genetic take-over, où il expose
l'idée d'une relève génétique, l'une des lois dont je vous ai parlé
tout au début. Dans ce livre, il imagine que nos ancêtres sont en fait
des ancêtres minéraux, que nous dérivons d'argiles qui se répliquaient
et qu'à partir de ces argiles, se sont constitués les systèmes vivants
tels qu'ils sont aujourd'hui. En fait, l'idée de Cairns-Smith, comme
vous allez le voir, contient deux éléments dont l'un est très
important et l'autre est inadéquat ; l'élément important est l'idée
d'un rôle important de la surface des solides. C'est ce que je vais
essayer de vous détailler par la suite. En revanche, l'idée inadéquate
était l'idée de se dire : si nous avons aujourd'hui un système qui se
reproduit selon une certaine hérédité, alors nous naissons d'un
système qui manifestait lui aussi une certaine hérédité. Cairns-Smith
cherchait donc systématiquement à prouver que les argiles peuvent se
répliquer. C'est évidemment très difficile. Les argiles sont, non pas
un ensemble linéaire comme un acide nucléique qui peut se reproduire
en copiant un fil sur un fil, mais à deux dimensions. Ainsi cela pose
déjà des problèmes assez compliqués, comment passer de deux dimensions
à une ? Ensuite, le grand nombre des expériences qu'il a pu faire ont
été assez ambiguës et n'ont pas vraiment donné les résultats
escomptés. Et je crois qu'une des raisons est simple, il aurait dû
pousser un peu plus son idée et penser non seulement à l'idée de
relève génétique, mais se dire qu'après tout, on n'a pas besoin de
génétique à l'origine.
Ce
que montre cette liste, c'est que, dans un mélange prébiotique, avec
une atmosphère imaginaire, on trouve un grand nombre de molécules plus
ou moins orientées. Et parmi ces molécules, quatre seulement sont des
acides aminés qui participent aux protéines aujourd'hui. Ainsi, l'un
des points tout-à-fait clair qui dérive de l'idée de bouillon
prébiotique, c'est qu'en fait on a un bouillon empoisonné, beaucoup
trop riche, mais dans lequel il manque des ingrédients essentiels.
C'est à ce stade qu'il devient intéressant de se demander comment on
pourrait imaginer à la fois un système qui puisse engendrer de la
variété mais surtout, introduire une sélection. C'est un point
essentiel sur lequel j'insiste parce que l'idée de sélection est une
idée très extrêmement répandue dans le monde anglo-saxon, mais
curieusement, très peu répandue en France où nous avons une tradition
assez particulière dans ce domaine. Quelle serait la sélection, le tri
assez efficace, pour ne retenir que certaines classes de
molécules ? Et ce sont ces molécules qui, elles, doivent pouvoir
constituer le premier métabolisme et engendrer un certain nombre des
éléments de base qui permettront la synthèse des êtres vivants. A ce
propos, un Anglais, Graham Cairns-Smith, depuis assez longtemps,
reprenant des idées antérieures, a eu une réflexion extrêmement
intéressante sur le rôle possible des argiles dans l'origine de la
vie. Il a écrit un livre, Genetic take-over, où il expose
l'idée d'une relève génétique, l'une des lois dont je vous ai parlé
tout au début. Dans ce livre, il imagine que nos ancêtres sont en fait
des ancêtres minéraux, que nous dérivons d'argiles qui se répliquaient
et qu'à partir de ces argiles, se sont constitués les systèmes vivants
tels qu'ils sont aujourd'hui. En fait, l'idée de Cairns-Smith, comme
vous allez le voir, contient deux éléments dont l'un est très
important et l'autre est inadéquat ; l'élément important est l'idée
d'un rôle important de la surface des solides. C'est ce que je vais
essayer de vous détailler par la suite. En revanche, l'idée inadéquate
était l'idée de se dire : si nous avons aujourd'hui un système qui se
reproduit selon une certaine hérédité, alors nous naissons d'un
système qui manifestait lui aussi une certaine hérédité. Cairns-Smith
cherchait donc systématiquement à prouver que les argiles peuvent se
répliquer. C'est évidemment très difficile. Les argiles sont, non pas
un ensemble linéaire comme un acide nucléique qui peut se reproduire
en copiant un fil sur un fil, mais à deux dimensions. Ainsi cela pose
déjà des problèmes assez compliqués, comment passer de deux dimensions
à une ? Ensuite, le grand nombre des expériences qu'il a pu faire ont
été assez ambiguës et n'ont pas vraiment donné les résultats
escomptés. Et je crois qu'une des raisons est simple, il aurait dû
pousser un peu plus son idée et penser non seulement à l'idée de
relève génétique, mais se dire qu'après tout, on n'a pas besoin de
génétique à l'origine.
Ce que je vais essayer de retenir maintenant, c'est l'idée d'un support solide, sans génétique au début, c'est-à-dire un système dans lequel on ne recherche pas un mécanisme héréditaire réplicatif initialement. Cette idée s'est développée dans plusieurs endroits dans le monde, en particulier mon jeune collègue, Philippe Marlière, avait eu un peu ce genre d'idées, mais surtout, très récemment, un Allemand qui travaille au Bureau des Brevets de Münich — j'ai trouvé que c'était assez drôle, en pensant à quelqu'un d'autre qui avait travaillé dans un bureau de brevets — a produit une théorie qui, à mon sens, est extrêmement remarquable. Ce monsieur s'appelle Günther Wächtershäuser, et sa théorie se résume ainsi : l'origine vient d'un métabolisme de surface, à la surface des solides que l'on trouve sur la terre, surface bien sûr en interface avec l'eau. Wächtershäuser a donné un grand nombre d'arguments pour justifier son point de vue. Son idée principale est que la sélection se fait par la charge électrique. C'est un point qui est très astucieux : si l'on regarde aujourd'hui — et je vais vous le montrer dans un instant — un tableau métabolique, on s'aperçoit qu'aucun chimiste, jamais, ne construirait les métabolites selon les modes de construction qu'on trouve dans un tableau métabolique. Ce serait complètement absurde. D'abord, cela se passe dans l'eau, et les chimistes n'aiment pas beaucoup travailler dans l'eau, comme vous le savez, ils travaillent plutôt dans des essences variées ; l'eau est un solvant particulièrement difficile pour la chimie ; et deuxièmement, ce sont des molécules subissant des transformations vraiment très, très bizarres. L'une de leurs bizarreries est que, très souvent, presque toujours en fait, elles portent à une de leurs extrémités, soit un groupe phosphate (qui est un groupe très fortement chargé négativement), soit un ou plusieurs groupes carboxylates (qui sont aussi chargés négativement). C'est ce point qu'a remarqué avec intérêt Wächtershäuser ; et il s'est dit que peut-être, la première sélection était que, justement, on ne retenait les molécules carbonées qu'à condition qu'elles possèdent des charges négatives leur permettant de rester en surface. Dans ce cas-là, vous voyez qu'on résout déjà un petit problème que je n'ai pas mentionné tout-à-l'heure à propos de la soupe prébiotique, le problème de la concentration. Le problème de concentration disparaît puisque tout se passe en surface : la surface concentre par elle-même ; dans un océan, dès qu'une molécule chargée a quelques chances de trouver la surface, même au bout d'un temps assez long, elle va s'y concentrer parce qu'elle restera retenue au voisinage de la surface. Essentiellement, les argiles, et toutes sortes de surfaces comme la pyrite de fer, sont des matrices chargées négativement porteuses d'ions métalliques positifs. Ceux-ci permettent de rejeter à la surface toute une série de charges positives susceptibles de retenir des molécules à la surface. Je ne vais pas, bien sûr, détailler ici les réactions chimiques que cela représente, je vais juste vous commenter très brièvement le tableau métabolique.
Voici un tout petit exemple qui
représente un tout petit morceau de tableau métabolique. Il s'agit
d'un morceau du chemin qui permet la dégradation du glucose, et la
synthèse de la sérine. Pourquoi vous montrer cela ? Parce qu'on y voit
des molécules à trois atomes de carbone, dans des réactions où se
passent des choses qui ne sont finalement pas très compliquées. 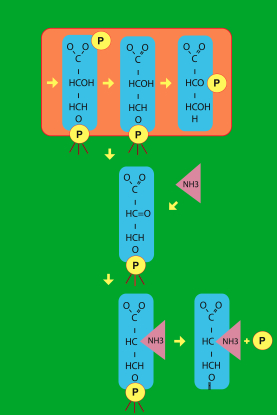 J'ai
mis sur la figure des petites "pattes" aux groupes phosphate pour
laisser imaginer qu'ils seraient effectivement liés à une surface. Il
faut donc considérer des groupes phosphate. Mais ici, dans l'exemple
choisi, je vous ai noté un cas tout-à-fait "absurde" du point de vue
utilitaire de la chimie en laboratoire et, en particulier, "absurde"
du point de vue énergétique. Et c'est là un point essentiel qu'il
convient aussi de retenir à propos de la vie : la vie ne se soucie pas
de perdre de l'énergie, elle préfère toujours perdre de l'énergie
plutôt que du contrôle. Cela vient du fait que ce qui compte pour la
vie, ce sont les relations entre les objets, et non les objets
eux-mêmes. Nous voyons ici la biosynthèse de cet acide aminé sérine.
Elle dérive directement de ces métabolites à trois carbones. Or ces
métabolites à trois carbones sont justement ceux à partir desquels
Wächtershäuser a construit toute son hypothèse. Et la particularité
chimique de cette hypothèse est qu'elle nécessite simplement une
transamination qui permet de récupérer l'azote pour faire l'acide
aminé. Je ne vais pas vous expliquer comment cela se réalise. Mais on
constate ici que, pour le faire, on doit passer par une sérine-phosphate,
qui possède donc un groupe phosphate. Or, ensuite, on doit enlever
le groupe phosphate pour fabriquer l'acide aminé sérine. C'est une
absurdité en apparence parce qu'on sait que, dans la cellule telle
qu'elle est aujourd'hui, ajouter des groupes phosphate coûte
énormément d'énergie. Il semble donc complètement absurde de vouloir
obtenir la sérine en passant par une étape contenant un phosphate pour
l'enlever ensuite... Il s'agit donc typiquement d'une trace du
métabolisme ancestral. Elle est restée figée parce que réinventer
quelque chose est extrêmement difficile lorsqu'on a déjà une solution
même coûteuse. On pourrait prendre comme analogie tout simplement
l'idée de la circulation sur une route. Vous savez tous que jusqu'à
Napoléon, on circulait à gauche, parce que nous sommes droitiers. Etre
droitier impose que nous ayions l'épée à gauche, et pour se battre ou
se défendre, il fallait être du côté gauche. Seul Napoléon, parce
qu'il avait imposé un système hiérarchisé extrêmement autoritaire, et
avec une idée particulière, extérieure au système lui-même, a pu
changer le côté de la circulation. Et l'on sait très bien combien il a
été très difficile, notamment en Suède, de le faire. Ce que je veux
dire c'est que si l'on s'est trouvé piégé dans un système métabolique
particulier, il peut être difficile ou impossible de revenir en
arrière, même s'il coûte de l'énergie. Et donc, pour moi, et c'est une
note d'espoir intéressante pour les chercheurs, cela veut dire qu'on
peut espérer trouver dans les êtres vivants aujourd'hui la trace du
passé (si l'on sait bien regarder).(4)
J'ai
mis sur la figure des petites "pattes" aux groupes phosphate pour
laisser imaginer qu'ils seraient effectivement liés à une surface. Il
faut donc considérer des groupes phosphate. Mais ici, dans l'exemple
choisi, je vous ai noté un cas tout-à-fait "absurde" du point de vue
utilitaire de la chimie en laboratoire et, en particulier, "absurde"
du point de vue énergétique. Et c'est là un point essentiel qu'il
convient aussi de retenir à propos de la vie : la vie ne se soucie pas
de perdre de l'énergie, elle préfère toujours perdre de l'énergie
plutôt que du contrôle. Cela vient du fait que ce qui compte pour la
vie, ce sont les relations entre les objets, et non les objets
eux-mêmes. Nous voyons ici la biosynthèse de cet acide aminé sérine.
Elle dérive directement de ces métabolites à trois carbones. Or ces
métabolites à trois carbones sont justement ceux à partir desquels
Wächtershäuser a construit toute son hypothèse. Et la particularité
chimique de cette hypothèse est qu'elle nécessite simplement une
transamination qui permet de récupérer l'azote pour faire l'acide
aminé. Je ne vais pas vous expliquer comment cela se réalise. Mais on
constate ici que, pour le faire, on doit passer par une sérine-phosphate,
qui possède donc un groupe phosphate. Or, ensuite, on doit enlever
le groupe phosphate pour fabriquer l'acide aminé sérine. C'est une
absurdité en apparence parce qu'on sait que, dans la cellule telle
qu'elle est aujourd'hui, ajouter des groupes phosphate coûte
énormément d'énergie. Il semble donc complètement absurde de vouloir
obtenir la sérine en passant par une étape contenant un phosphate pour
l'enlever ensuite... Il s'agit donc typiquement d'une trace du
métabolisme ancestral. Elle est restée figée parce que réinventer
quelque chose est extrêmement difficile lorsqu'on a déjà une solution
même coûteuse. On pourrait prendre comme analogie tout simplement
l'idée de la circulation sur une route. Vous savez tous que jusqu'à
Napoléon, on circulait à gauche, parce que nous sommes droitiers. Etre
droitier impose que nous ayions l'épée à gauche, et pour se battre ou
se défendre, il fallait être du côté gauche. Seul Napoléon, parce
qu'il avait imposé un système hiérarchisé extrêmement autoritaire, et
avec une idée particulière, extérieure au système lui-même, a pu
changer le côté de la circulation. Et l'on sait très bien combien il a
été très difficile, notamment en Suède, de le faire. Ce que je veux
dire c'est que si l'on s'est trouvé piégé dans un système métabolique
particulier, il peut être difficile ou impossible de revenir en
arrière, même s'il coûte de l'énergie. Et donc, pour moi, et c'est une
note d'espoir intéressante pour les chercheurs, cela veut dire qu'on
peut espérer trouver dans les êtres vivants aujourd'hui la trace du
passé (si l'on sait bien regarder).(4)
Pour en revenir aux pierres, il y a un deuxième aspect qui est très important que je ne peux pas discuter longtemps, faute de temps, c'est le rôle du deuxième principe de la thermodynamique dans le système vivant. On a tendance, à la suite de Bolzmann au siècle dernier, à faire comme si le deuxième principe de la thermodynamique était une contrainte contre laquelle nous devons lutter. C'est tout-à-fait inexact, le deuxième principe est tout-à-fait normal, c'est un principe physique absolument habituel avec lequel la vie compose comme avec tous les principes physiques. En particulier, ici, dans le cas précis de ce qui se passe quand on considère un système à deux dimensions (sur une surface) et que l'on compare à ce qui se passe à trois dimensions (dans un liquide), eh bien, le deuxième principe de la thermodynamique dit la chose suivante : en surface, la polymérisation est favorisée et c'est là un point extrêmement important, alors qu'à l'inverse, dans un liquide, la dépolymérisation est favorisée. En appliquant le deuxième principe et, en particulier, parce que les polymérisations dans les systèmes vivants sont des polymérisations qui se font avec élimination d'une molécule d'eau, on comprend très bien ce qui se passe. En surface, on trouve d'abord des molécules qui restent chargées ; elles peuvent diffuser, s'orienter les unes par rapport aux autres, mais le nombre de leurs degrés de liberté n'est finalement pas très grand puisqu'elles restent en surface. Mais si elles se combinent, elles éliminent une molécule d'eau. Or, justement, cette molécule d'eau n'est pas chargée, elle peut donc partir facilement dans la solution. Elle a par conséquent un très grand nombre de degrés de liberté. Ainsi, la surface favorise la polymérisation. Alors, si l'on considère à nouveau, non pas le bouillon de culture, mais la surface, l'idée de la genèse de polymères, dont on va avoir besoin puisqu'il va falloir édifier des protéines des acides nucléiques, est une conséquence toute naturelle du métabolisme de surface.
Un deuxième point que je ne vais pas détailler ici, c'est qu'on peut montrer, tout simplement à partir de l'observation du métabolisme aujourd'hui, que certains lipides pourraient parfaitement être synthétisés en surface. C'est à nouveau une conséquence du rôle de la surface. Comme je viens de vous le montrer, à propos de l'acide aminé sérine, on constate une bizarrerie chimique dans la synthèse d'un grand nombre de lipides (composants de base des membranes). Cette bizarrerie est la suivante : ils dérivent de molécules qui contiennent un groupe pyrophosphate, c'est-à-dire deux phosphates à l'une de leurs extrémités. Là aussi, cela paraît être au premier abord une absurdité. A priori, il n'y a aucune raison pour expliquer ce fait, si ce n'est à nouveau la trace d'une histoire. C'est donc l'idée de Wächtershaüser, qu'il détaille très longuement dans son article. Les lipides, ou du moins, certaines classes de lipides, ceux qu'on trouve aujourd'hui chez les bactéries qu'on appelle les archébactéries, ces lipides peuvent avoir été fabriqués dans une phase tout-à-fait précoce (5). Cela nous amène donc à une situation qui a une certaine originalité, puisqu'elle permet de sélectionner en surface. Ainsi s'élabore une certaine variété de molécules et parmi cette variété, on peut expliquer et la synthèse de certains lipides à partir desquels se forme une membrane et, aussi, ce qui mériterait d'être détaillé, la synthèse de certains co-enzymes. En particulier, la sérine pour laquelle je vous ai montré tout-à-l'heure un petit morceau de chemin métabolique, est très apparentée, par sa biosynthèse, à celle d'un co-enzyme, c'est-à-dire d'un facteur qui sert à faire des réactions enzymatiques (dont celle qui produit la sérine elle-même). Donc, on s'aperçoit qu'en surface sont réunis les ingrédients permettant de faire la catalyse, de la favoriser et de faire la polymérisation des métabolites élémentaires et même des nucléotides. Nous sommes, si l'on choisit l'hypothèse d'un métabolisme de surface, dans une situation extrêmement prometteuse.
Toute la question va être alors d'imaginer ce qui a pu se produire dans le détail. A ce propos, je voudrais juste vous mentionner ce que cela signifie concrètement. Comment va-t-on pouvoir regarder tous ces processus ? C'est évidemment à partir des êtres vivant aujourd'hui qu'il faudra commencer, et c'est là-dessus que je terminerai tout-à-l'heure cet exposé. On doit donc tenter de distinguer dans les êtres vivant aujourd'hui ce qui peut revenir à ce qui est ancestral et ce qui, au contraire, est nouveau. Nous venons de voir que si tout a commencé en surface, cela va conduire assez rapidement à la synthèse d'une membrane, des lipides qui se font autour des particules solides. On crée donc de façon irréversible, et de façon tout-à-fait émergente, un objet nouveau. Une membrane qui se referme avec une propriété aussi simple (c'est, notons-le, un nouveau problème de relation entre objets) que le fait de se refermer crée un intérieur et un extérieur. Or cela, c'est irréductible à tout ce qui existait auparavant ; dès qu'on est dans cette situation, le métabolisme se trouve emprisonné à l'intérieur de la membrane. S'il est emprisonné à l'intérieur de la membrane, la nécessité d'un support solide, ainsi que des attaches à ce support disparaît. On s'attend donc à ce que la dégradation des objets — pour l'instant, je vous ai parlé surtout de leur construction, c'est-à-dire ce qu'on appelle l'anabolisme — la partie au contraire destructive, celle qui consiste à récupérer les choses dans l'environnement, à partir des premières cellules. n'a aucune raison d'être de même nature. Et effectivement, si l'on regarde soigneusement les tableaux métaboliques, c'est-à-dire l'ensemble des règles qui font correspondre les différents produits les uns aux autres, on s'aperçoit que ce qui est anabolique contient des groupements chimiques chargés, en particulier des phosphates, alors que ce qui est catabolique ne les contient plus. Là encore, nous avons l'impression qu'aujourd'hui, nous sommes dans une situation qui, après tout, n'est peut-être pas si éloignée des origines que cela.
Pour l'instant, nous nous trouvons
donc simplement en présence d'un métabolisme de surface, donc de
fonctions, mais n'ont pas encore été réellement fabriquées les
macromolécules qui vont être essentielles. Et je ne vous ai rien dit,
bien sûr, de la synthèse des nucléotides ou d'autres molécules de ce
genre. Nous nous trouvons donc amenés à nous poser des questions sur
la façon dont se produit ce métabolisme de surface et surtout, sur la
façon dont ensuite il va être transformé à l'intérieur de la cellule.
Un concept particulier que j'appelle la transformation homéotopique
permet d'y répondre : on se trouve en présence d'une surface comme je
l'ai présentée, puis de molécules dont l'une peut être acceptrice,
(ici, c'est celle qui est représentée avec un rectangle) et l'autre
donneuse du groupement chimique, un co-enzyme, par exemple, est ce qui
donne et qui transfère d'un objet à l'autre.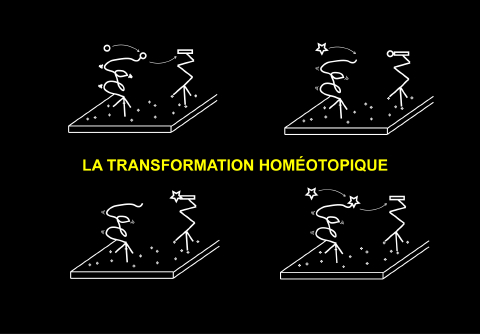 Ensuite, si je continue mon schéma, j'ai représenté de façon un peu
cinématographique par des petits ronds, puis un triangle, un deuxième
type de molécules, obtenu à partir du même substrat et d'un co-enzyme
qui peut transférer des objets diffférents. Il s'agit là d'une
transformation typique du métabolisme d'aujourd'hui. Tout le problème
va donc être d'imaginer quels seront les objets qui auront pu se
substituer à la surface pour permettre ce genre de transformation
homéotopique. Dit autrement, est-ce qu'il existe des molécules
acceptrices sur lesquelles on peut faire des variations, et à partir
de ces variations, obtenir les nouvelles molécules qui seront
utilisées pour les synthèses suivantes ?
Ensuite, si je continue mon schéma, j'ai représenté de façon un peu
cinématographique par des petits ronds, puis un triangle, un deuxième
type de molécules, obtenu à partir du même substrat et d'un co-enzyme
qui peut transférer des objets diffférents. Il s'agit là d'une
transformation typique du métabolisme d'aujourd'hui. Tout le problème
va donc être d'imaginer quels seront les objets qui auront pu se
substituer à la surface pour permettre ce genre de transformation
homéotopique. Dit autrement, est-ce qu'il existe des molécules
acceptrices sur lesquelles on peut faire des variations, et à partir
de ces variations, obtenir les nouvelles molécules qui seront
utilisées pour les synthèses suivantes ?
Ici, je vais prendre un raccourci, faire comme si l'on avait résolu le problème de l'existence des nucléotides. C'est à mon sens, actuellement le problème qui, de très loin, est le plus difficile. Et je ne cache pas que jusqu'au bout de mon exposé, cela restera un peu un mystère. Je vous proposerai quand même une hypothèse assez amusante à ce propos, mais cela reste quelque chose qu'on peut considérer comme une question majeure. Vous avez certainement entendu parler récemment des ARN catalytiques — il y a eu cette année deux prix Nobel à ce sujet. Il s'agit d'une découverte extrêmement intéressante. Je vous ai parlé tout-à-l'heure de mémoire et de fonction, c'est-à-dire des acides nucléiques qui jouaient le rôle de mémoire et des protéines qui étaient manipulatrices, qui possédaient la fonction. Tom Cech a découvert en 1981 que certains ARN pouvaient avoir les deux activités, c'est-à-dire pouvaient à la fois se reproduire eux-mêmes et avoir des activités catalytiques. Cela résout évidement en partie le paradoxe de l'œuf et de la poule, qui est le premier, acide nucléique ou protéine ? Les ARN sont premiers, ils réalisent les deux à la fois. Je vous ferai remarquer, cependant, que si les gens ont beaucoup crié cocorico à ce propos — c'est le cas de le dire à propos de l'œuf et de la poule — c'était un peu mal venu, car comme je vous l'ai dit, la question de l'origine des nucléotides nécessaires à la synthèse des ARN reste parfaitement ouverte. Néanmoins, il est vrai qu'à partir du moment où l'on admet — et je pense que c'est tout-à-fait admissible — que les ARN ont été inventés, alors, on comprend très bien la suite de l'évolution. En effet, la suite va être cruciale, elle va substituer à un objet qui a deux particularités, celle de transmettre la mémoire et celle de réaliser une fonction, deux objets permettant de séparer mémoire et fonction. Et c'est cela qui permettra de créer le codage et qui permettra de créer la vie. La vie n'est pas encore créée avec les ARN, il s'en faut de beaucoup. Ce qui est essentiel dans la vie, comme je vous l'ai dit au début, c'est cette relation tout-à-fait originale qui transpose une mémoire et une fonction. C'est cela qui permet au système de prendre une distance par rapport à lui-même et de faire, avec le deuxième code que nous utilisons aujourd'hui, le langage, de faire ce que nous sommes en train de faire, de nous retourner sur nous-mêmes. Mais la découverte des ARN catalytiques est effectivement un très grand pas dans la compréhension de l'origine de la vie. L'idée qu'on peut avoir pour expliquer leur apparition, c'est que ces ARN catalytiques ont en fait d'abord joué le rôle de substitut des surfaces minérales. Pourquoi peut-on penser cela ? Les ARN sont des polyanions extrêmement chargés, ils ont des groupes phosphate partout, ce qui leur permet de lier des ions positifs. Donc, ils jouent un peu le même rôle que les surfaces que j'ai mentionnées tout-à-l'heure. Ainsi, la question qu'on peut se poser, est : ne sont-ils pas eux-mêmes soumis à des règles métaboliques ? Or, il existe parmi ces ARN une catégorie particulière qu'on appelle les ARN de transfert. Il s'agit des objets qui font la correspondance entre la mémoire et la fonction. Ce sont eux qui font la correspondance entre ce qui est écrit dans le texte des nucléotides et la protéine. Ces ARN de transfert ont deux particularités, d'une part leur taille, ce sont d'assez grosses molécules dotées d'environ une centaine de nucléotides, aussi en tenant compte de la diffusion des réactions chimiques habituelles, ils bougent lentement par rapport à la diffusion des petites molécules, ce qui leur permet de jouer le rôle de support ; et d'autre part, et c'est, à mon sens, extrêmement important, ils sont eux-mêmes très fortement modifiés métaboliquement. On sait en effet que les ARN de transfert, au lieu d'être une suite de nucléotides classiques, contiennent au moins un quart de nucléotides totalement modifiés. Et dans un très grand nombre de cas, on n'a pas la moindre idée de la raison d'être de ces modifications. Aussi, peut-on se demander si ces modifications n'interviennent pas dans le métabolisme. On sait aussi, et cela apparaît maintenant de plus en plus souvent, que les ARN de transfert interviennent dans un grand nombre de réactions qui n'ont rien à voir avec leur rôle d'ARN de transfert, c'est-à-dire de faire passer le message de l'ARN messager à la protéine.
Alors, donc, pour schématiser un peu ce
que je viens de vous dire, on peut imaginer que les transformations de
type homéotopique, comme je vous les ai décrites tout-à-l'heure, se
déroulent sur des ARN de transfert. Alors, au lieu de représenter une
surface, on peut représenter un ARN de transfert porteur de réactions
de ce type. Et je dois vous dire que l'une des raisons pour lesquelles
je suis ici ce soir, c'est que nous avons eu la chance de découvrir
dans mon laboratoire, accidentellement, une transformation de ce type
qui suggère que, en effet, les ARN de transfert ont joué ce rôle. Nous
avons trouvé, en comparant deux protéines qu'elles étaient visiblement
apparentées et servaient à faire la synthèse de deux acides aminés qui
n'avaient aucun rapport entre eux excepté le fait que leur substrat
initial, ici représenté par un rectangle, était le même, mais qu'il
subissait une modification de type homéotopique.  Et
quand on expose un peu plus cette réaction, on s'aperçoit que le
substrat — le rectangle — est l'acide aminé sérine, dont je vous ai
parlé. Or, on le retrouve dans un certain nombre de cas, et en
particulier, dans la synthèse d'un vingt et unième acide aminé que je
vous ai mentionné au début, découvert très récemment comme étant
incorporé dans certaines protéines. Dans ce dernier cas, on remarque
que la modification se fait de la façon suivante : l'ARN de transfert
tient comme une pince-étau un premier acide aminé (la sérine, souvent)
et cet acide aminé est modifié par une activité enzymatique qui lui
rajoute un groupement chimique ou un autre. Par exemple, il est chargé
par la sérine, et dans le cas du vingt et unième acide aminé, la
sélénocystéine, il lui est ajouté du sélénium. En réalité, quand on
regarde de près, on s'aperçoit qu'il existe un grand nombre de
réactions dans lesquelles les ARN de transfert sont impliqués, et où
ils servent de support à une molécule du métabolisme modifiée in
situ sur l'ARN de transfert. C'est le cas, par exemple, de la
méthionine qui peut être modifiée par un groupe formyl. C'est aussi le
cas d'un acide aminé tout-à-fait important, la glutamine, qui, chez
beaucoup d'organismes, en particulier, chez les bactéries Gram
positives, au lieu d'être chargé directement sur son ARN de transfert
comme on l'attend dans la correspondance du code génétique, est
modifié à partir de l'acide glutamique. Ainsi, ce n'est pas le bon
acide aminé qui est chargé, mais un autre et par modification
homéotopique, il est transformé en celui qu'il faudrait. Tout se passe
comme si l'ARN de transfert jouait le rôle de support d'une molécule
qui sera modifiée par la suite. Par ailleurs, on a découvert que des
ARN de transfert interviennent directement dans la synthèse des parois
bactériennes ou dans la synthèse d'un co-enzyme comme la chlorophylle.
Pour faire de la chlorophylle, il existe une étape où un ARN de
transfert tient un résidu glutamate mais cet ARN de transfert n'a en
principe aucune fonction dans la synthèse elle-même. On se rend compte
alors que peut-être, ce sont là encore des traces de l'histoire
évolutive parce qu'on n'a absolument pas besoin de cet ARN de
transfert pour faire la réaction chimique en question. D'ailleurs,
dans le cas de la chlorophylle — il s'agit de la synthèse d'un
métabolisme précurseur qui s'appelle l'amino-lévulinate, il existe en
fait dans la nature deux voies de synthèse différentes ; une voie chez
les bactéries utilise l'ARN de transfert chargé par la glutamate mais
chez les mammifères par exemple, le rôle de cet ARN a disparu.
Lorsqu'on observe son existence, il s'agit donc d'un reste du passé,
d'une archive oubliée. Je ne vais retenir qu'un point de cette petite
digression, parce qu'elle va me permettre de revenir aux nucléotides
comme je vous l'ai promis, je vais retenir le rôle de la synthèse
peptidique.
Et
quand on expose un peu plus cette réaction, on s'aperçoit que le
substrat — le rectangle — est l'acide aminé sérine, dont je vous ai
parlé. Or, on le retrouve dans un certain nombre de cas, et en
particulier, dans la synthèse d'un vingt et unième acide aminé que je
vous ai mentionné au début, découvert très récemment comme étant
incorporé dans certaines protéines. Dans ce dernier cas, on remarque
que la modification se fait de la façon suivante : l'ARN de transfert
tient comme une pince-étau un premier acide aminé (la sérine, souvent)
et cet acide aminé est modifié par une activité enzymatique qui lui
rajoute un groupement chimique ou un autre. Par exemple, il est chargé
par la sérine, et dans le cas du vingt et unième acide aminé, la
sélénocystéine, il lui est ajouté du sélénium. En réalité, quand on
regarde de près, on s'aperçoit qu'il existe un grand nombre de
réactions dans lesquelles les ARN de transfert sont impliqués, et où
ils servent de support à une molécule du métabolisme modifiée in
situ sur l'ARN de transfert. C'est le cas, par exemple, de la
méthionine qui peut être modifiée par un groupe formyl. C'est aussi le
cas d'un acide aminé tout-à-fait important, la glutamine, qui, chez
beaucoup d'organismes, en particulier, chez les bactéries Gram
positives, au lieu d'être chargé directement sur son ARN de transfert
comme on l'attend dans la correspondance du code génétique, est
modifié à partir de l'acide glutamique. Ainsi, ce n'est pas le bon
acide aminé qui est chargé, mais un autre et par modification
homéotopique, il est transformé en celui qu'il faudrait. Tout se passe
comme si l'ARN de transfert jouait le rôle de support d'une molécule
qui sera modifiée par la suite. Par ailleurs, on a découvert que des
ARN de transfert interviennent directement dans la synthèse des parois
bactériennes ou dans la synthèse d'un co-enzyme comme la chlorophylle.
Pour faire de la chlorophylle, il existe une étape où un ARN de
transfert tient un résidu glutamate mais cet ARN de transfert n'a en
principe aucune fonction dans la synthèse elle-même. On se rend compte
alors que peut-être, ce sont là encore des traces de l'histoire
évolutive parce qu'on n'a absolument pas besoin de cet ARN de
transfert pour faire la réaction chimique en question. D'ailleurs,
dans le cas de la chlorophylle — il s'agit de la synthèse d'un
métabolisme précurseur qui s'appelle l'amino-lévulinate, il existe en
fait dans la nature deux voies de synthèse différentes ; une voie chez
les bactéries utilise l'ARN de transfert chargé par la glutamate mais
chez les mammifères par exemple, le rôle de cet ARN a disparu.
Lorsqu'on observe son existence, il s'agit donc d'un reste du passé,
d'une archive oubliée. Je ne vais retenir qu'un point de cette petite
digression, parce qu'elle va me permettre de revenir aux nucléotides
comme je vous l'ai promis, je vais retenir le rôle de la synthèse
peptidique.
Certains ARN de transfert interviennent donc pour transférer leur acide aminé de l'un à l'autre en formant un lien peptidique. Or, faire un lien peptidique, c'est en fait commencer à fabriquer une protéine, c'est exactement ainsi que sont synthétisées les protéines aujourd'hui. Mais les ARN de transfert ont alors une deuxième fonction, celle d'assurer la correspondance entre un ARN, l'ARN messager et la suite des acides aminés dans la protéine. C'est ce qui assure les liens entre la mémoire (l'acide nucléique) et la fonction (la protéine) en obéissant à la règle du code génétique. Cependant, dans certains cas, certaines protéines ont un acide aminé rajouté à une de leur extrémité par un ARN de transfert, mais en dehors du contexte de l'expression du message génétique. Cela conduit donc à penser que les ARN de transfert pouvaient peut-être, à l'origine, avoir deux fonctions. L'une les associait via des ARN autocatalytiques au mécanisme de leur propre reproduction. L'autre les voyait supports du métabolisme, avec pour fonction, entre autres choses, de fabriquer des peptides. L'idée, ici, est qu'il est possible d'imaginer que très tôt, et peut-être même avant les nucléotides, comme je vous l'ai dit, des peptides existaient. C'est là un point assez important, qui renouvelle le problème de l'œuf et de la poule. A nouveau, ce sont les protéines qui vont être antérieures, mais cette fois-ci, sous la forme de petits peptides. Les premiers ARN de transfert auraient pu servir de supports pour la fabrication de peptides et c'est au cours de cette activité qu'ils ont découvert, si l'on peut dire, que le fait de s'aligner séquentiellement de façon appropriée, leur permettait d'avoir une plus grande efficacité dans la fabrication des peptides. Or, un moyen d'aligner des acides nucléiques, étant donnée leur capacité à créer des appariements par complémentarité comme dans la double hélice d'ADN, est de former ceux qu'on découvre dans la lecture du messager. On voit là comment pourrait avoir été inventée la lecture d'un messager initial comme favorisant la fabrication de peptides. Donc, la traduction telle qu'on la connaît aujourd'hui, c'est-à-dire le passage des ARN messagers vers les protéines, serait une retombée de l'évolution vers une plus grande spécificité dans la fabrication des peptides. Or, si l'on arrive à cela, on crée la séparation entre la mémoire et la fonction, puisque pour faire certains peptides, certaines classes d'ARN de transfert seront préférées, ce qui crée de facto une correspondance entre certains ARN supports (ce seront les messagers) et certaines protéines.
Mais revenons à la question "est-il possible d'imaginer la genèse des nucléotides ?" La littérature actuelle expose partout la fabrication des purines. Elle met toujours en avant de l'acide cyanhydrique, de nombreux processus qui se déroulent dans les étoiles. Mais c'est bien peu plausible, d'autant plus qu'il manque les pyrimidines tout aussi nécessaires et que si l'on considère les purines, on en fabrique toutes sortes qui ne sont pas les bonnes. Or, si l'on regarde les tableaux métaboliques actuels, on s'aperçoit que les bases nucléiques sont fabriquées à partir de sucres phosphorylés — donc typiquement des molécules associées à un support minéral, comme je l'ai dit tout-à-l'heure — et d'acides aminés. C'est là un point absolument essentiel, aujourd'hui, dans le métabolisme actuel, on commence à prendre un phosphoribosyl pyrophosphate sur lequel une glutamine (un acide aminé par conséquent) transfère un groupe aminé. C'est la première étape. Il est donc légitime de penser que la genèse de certains co-enzymes et des bases nucléiques s'est faite initialement à partir de petits peptides. Ainsi, c'est la synthèse, la genèse de la liaison peptidique qui va être le coeur des premières étapes. Ensuite, comme je l'ai dit, une fois qu'ont été formés les premiers nucléotides, ils vont avoir tendance à polymériser, à édifier des ARN de transfert et, par conséquent, à favoriser précisément la fabrication des peptides qui permettent eux-mêmes de former les bases des acides nucléiques. On a là un système autogène, qui a l'aptitude à favoriser sa propre création, à partir de peptides. Or, il existe aujourd'hui un système dans lequel on connaît la genèse de peptides précis en l'absence d'acides nucléiques, mais à l'aide de peptides. C'est une observation que je trouve assez intéressante : il existe parmi les différents co-enzymes un peptide phosphorylé (le 4-phosphopantéthéine) qui se comporte essentiellement comme un bras articulé porteur d'un soufre à une extrémité. Cet atome est assez actif et permet de transporter grâce au bras divers groupes chimiques d'un endroit à un autre. Ce co-enzyme est impliqué dans la genèse des lipides. Mais on a découvert assez récemment, et c'est quelque chose qui me semble extrêmement frappant, qu'il intervient aussi dans la genèse de plusieurs antibiotiques peptidiques. Cela se fait de la façon suivante. Une structure cyclique faite de plusieurs protéines permet de lier plusieurs acides aminés et le bras va aller les chercher les uns après les autres et les enchaîner par une liaison peptidique. Or, cette structure est très fortement apparentée à celle qui permet de faire des lipides, les motifs enchaînés seuls diffèrent. On peut donc parfaitement imaginer que, dans une mécanique initiale, on ait un complexe de ce genre qui ait engendré de façon assez efficace à la fois la genèse de peptides et de lipides. Et il est ensuite légitime de penser qu'à partir de ces peptides, seront construits les nucléotides, ceux-là à leur tour joueront le rôle des substituts des surfaces et permettront ensuite la genèse des êtres vivants.
Est-il possible, aujourd'hui encore, de voir la trace de ce que je viens de décrire ? Bien sûr, dans les comparaisons entre les différents objets qu'on peut découvrir, cela peut se voir. Mais je crois que cela se voit aussi dans un ensemble particulier, le tableau du code génétique (que j'ai redessiné ici d'une façon hétérodoxe). Le tableau du code génétique est essentiellement la correspondance entre les lettres de l'alphabet des acides nucléiques et les vingt acides aminés.
|
Lys (K)
|
AAA
|
Arg (R)
|
AGA
|
Thr (T)
|
ACA
|
I/M
|
ATA
|
|
|
AAG
|
|
AGG
|
|
ACG
|
Met (M)
|
ATG
|
|
Asn (N)
|
AAC
|
Ser (S)
|
AGC
|
|
ACC
|
Ile (I)
|
ATC
|
|
|
AAT
|
|
AGT
|
|
ACT
|
|
ATT
|
|
Glu (E)
|
GAA
|
Gly (G)
|
GGA
|
Ala (A)
|
GCA
|
Val(V)
|
GTA
|
|
|
GAG
|
|
GGG
|
|
GCG
|
|
GTG
|
|
Asp (D)
|
GAC
|
|
GGC
|
|
GCC
|
|
GTC
|
|
|
GAT
|
|
GGT
|
|
GCT
|
|
GTT
|
|
Gln (Q)
|
CAA
|
Arg (R)
|
CGA
|
Pro (P)
|
CCA
|
Leu (L)
|
CTA
|
|
|
CAG
|
|
CGG
|
|
CCG
|
L/S
|
CTG
|
|
His (H)
|
CAC
|
|
CGC
|
|
CCC
|
Leu (L)
|
CTC
|
|
|
CAT
|
|
CGT
|
|
CCT
|
|
CTT
|
|
Fin (*)
|
TAA
|
W/Se/*
|
TGA
|
Ser (S)
|
TCA
|
Leu (L)
|
TTA
|
|
*/Q
|
TAG
|
Trp (W)
|
TGG
|
|
TCG
|
|
TTG
|
|
Tyr (Y)
|
TAC
|
Cys (C)
|
TGC
|
|
TCC
|
Phe(F)
|
TTC
|
|
|
TAT
|
|
TGT
|
|
TCT
|
|
TTT
|
|
|
|
|
|
|
|
|
|
| Dérivés des acides dicarboxyliques | ||
| Dérivés de la sérine-phosphate | ||
| Dérivés de l'homosérine-phosphate |
Ce qu'on voit ici, c'est qu'on peut relier la construction de ce code, que je ne vais pas détailler, à des règles du métabolisme. Et dans ces règles, interviendrait une molécule chargée, soit la sérine-phosphate, soit l'homosérine phosphate, soit les acides dicarboxyliques. On peut considérer, comme Wong, il y a quelques années, l'avait déjà proposé, que la genèse du code vient non pas, comme on l'a beaucoup recherchée, d'une adéquation entre les codons, c'est-à-dire les suites de nucléotides correspondant au message, et les acides aminés, mais de la trace de l'histoire métabolique qui a donné naissance aux acides aminés en question. C'est-à-dire que l'on retrouverait tout simplement l'idée de la transformation homéotopique par laquelle certains acides aminés pouvaient être liés à des ARN de transfert et modifiés. Et c'est ce processus métabolique qui aurait engendré les règles du code. Les codons apparentés pourraient avoir été dérivés d'ARN de transfert apparentés par ce métabolisme. Vous allez me dire que tout cela est très imaginaire : pourquoi et comment peut-on aujourd'hui remonter à l'origine ? Bien sûr, un certain nombre d'auteurs ont essayé de faire des expériences, comme je vous l'ai dit, à partir d'étincelles et d'autres sources d'énergie dans un mélange gazeux. Cela aussi supposait imaginer un scénario d'origine, imaginer des supports, imaginer une atmosphère d'origine, toutes questions qui sont évidemment extrêmement aléatoires. Mais il y a un autre moyen d'approche qui, à mon sens, est extrêmement puissant et qui est en train de se mettre en place, et c'est celui-là sur lequel je vais terminer. C'est d'ailleurs cette autre approche qui m'a amené à me poser des questions d'origines. Cet autre moyen d'approche est l'étude complète du génome des êtres vivants aujourd'hui. Vous avez sans doute entendu parler d'un de ces génomes, ce génome pèse environ deux cents millions de dollars aux Etats-Unis en ce moment, c'est le nôtre. Comme vous allez le voir immédiatement ici, c'est un projet fascinant du point de vue médiatique, mais qui soulève de nombreuses questions, en particulier pour de nombreuses raisons scientifiques, mais surtout pour des raisons éthiques. Ici, sont indiqués la plupart des projets correspondant à la détermination complète du texte du génome.
|
GENOME |
TOTAL |
GENES |
|
|
|
|
|
Homo sapiens |
3 10 9 |
50-100 103 |
|
Arabidopsis thaliana |
70 10 6 |
20-50 103 |
|
Drosophila melanogaster |
40 10 6 |
25 103 |
|
Saccharomyces cerevisiae |
17 10 6 |
12 103 |
|
Bacillus subtilis |
5 10 10 6 |
4.5 103 |
|
Escherichia coli |
4.7 10 6 |
4.5 103 |
|
Chlamydia trachomatis |
1.2-1.4 10 6 |
? 1.2 103 |
|
Mycoplasma sp. |
0.8-1.2 10 6 |
1 103 |
|
|
|
|
|
VIRUS |
|
|
|
|
|
|
|
DNA:Cytomegalovirus |
450 103 |
450 |
|
Variole |
200 103 |
200 |
|
RNA:SIDA |
15 103 |
15 |
|
Rage |
12 103 |
12 |
|
Phage R17 |
3 103 |
3 |
Vous voyez donc que, dans le cas de l'Homme, le génome est composé d'environ trois milliards de lettres. C'est énorme et il s'y ajoute une difficulté importante du fait qu'il n'existe pas de lignées humaines fortement consanguines, il existe un polymorphisme considérable : plus de 5%. Cela implique un problème considérable de contrôle interne dans l'analyse du génome. Et c'est vrai quelles que soient les circonstances. Une deuxième difficulté est que, comme je vous l'ai dit initialement, dans tout ce qui est vivant, ce qui compte essentiellement est le contrôle, beaucoup plus que l'énergie. Pour cette raison, les êtres vivants ne se soucient pas d'être chargés de fardeaux pourvu qu'ils ne soient pas trop lourds, dans la mesure où ils pourront servir un jour. C'est un point, je crois, tout-à-fait essentiel : on peut transporter avec soi des archives — nous le faisons d'ailleurs, nous avons des ministères entiers d'archives — dont on perd les pages de temps en temps, on les véhicule lors des déménagements, parce que peut-être, un jour, elles pourront servir. Typiquement, dans un génome humain, ce qui revient aux gènes qui sont réellement exprimés, correspond à peu près à 2% du génome. Vouloir déterminer d'emblée la séquence totale de l'ensemble pose donc des questions très difficiles, non seulement parce qu'on ne disposera pas d'un contrôle interne de fiabilité, mais parce qu'il sera très difficile de découvrir les gènes dans un océan d'archives. Savoir distinguer ce qui est pertinent de ce qui ne l'est pas paraît extrêmement difficile. Par ailleurs, ce qu'il faudrait, c'est pouvoir faire ce que nous appelons, nous généticiens, la génétique inverse, c'est-à-dire revenir en arrière en inactivant le gène présumé pour voir ce qui se passe. Dans ce cas précis, l'Homme est particulièrement mal adapté, à moins qu'on ait des idées spécialement mengéliennes sur l'humanité. On peut donc écarter l'étude du génome humain, du moins en tant que projet de séquençage, comme étant pertinent pour notre propos. En revanche, les choses sont tout-à-fait différentes pour toute une série d'autres organismes.
Et ce que je voudrais vous faire remarquer ici, c'est qu'on en arrive assez rapidement à l'étude d'êtres tout-à-fait autonomes comme les levures, les colibacilles, toutes sortes de microbes d'intérêt industriel ou médical et même d'autres organismes comme les mycoplasmes où l'on arrive à peu près à un million de lettres, ce qui est un texte extrêmement court. Par comparaison, le cytomégalovirus qui a été séquencé par un groupe en Angleterre fait le quart de cette séquence. Ainsi, vous voyez qu'on arrive à des conditions dans lesquelles on peut avoir accès à la détermination complète de la séquence d'un génome. Cependant, il ne faut pas croire que c'est un projet aisé en lisant les chiffres qui sont souvent indiqués dans la littérature et qui ont essentiellement un rôle médiatique ou commercial. Les choses sont beaucoup plus laborieuses que ce qui est dit. Actuellement, personne au monde ne sait déterminer sans erreur plus de 50 000 paires de bases par an et par personne (6). Vous voyez pourtant que pour un million de paires de bases, ce n'est pas gigantesque. Cela peut actuellement être envisagé avec des petites équipes, par rapport aux projets de la physique, par exemple. En revanche, ce qu'on obtient est uniquement un texte et là encore, malheureusement, vient une idée reçue qui est tout-à-fait mal venue, l'idée que l'on peut comparer ce texte à un message téléphonique à propos duquel on pourrait faire une analyse probabiliste, en fonction de la probabilité de présence de telle ou telle base ou acide aminé à telle ou telle position. Or, un gène n'a rien à voir avec un message téléphonique. Ce qu'il est essentiel de retenir, c'est qu'un ADN, c'est quelque chose qui a une histoire ; chaque base n'est pas un élément qui a une probabilité de se trouver là où elle est, c'est un élément qui a une histoire. Et c'est découvrir cette histoire qui est essentiel. On peut le faire à la fois à l'intérieur d'un même organisme, en comparant les différents gènes et les diférentes protéines et en créant les arbres généalogiques qui les relient les unes aux autres : et on peut aussi le faire en comparant les organismes entre eux. C'est en découvrant un de ces aspects comparatifs que j'ai été amené à me persuader de leur pertinence. On peut établir des comparaisons, découvrir des parentés entre des chemins métaboliques qui n'ont apparemment aucun rapport. Cela amène nécessairement à se poser la question de l'existence d'un ancêtre commun et donc à remonter vers les origines, et à comprendre que le rapport entre ces chemins métaboliques est un rapport d'histoire. En parallèle, on découvre un aspect tout-à-fait fabuleux : vous le savez sans doute, on peut, en comparant par exemple des protéines ayant des fonctions homologues dans des organismes différents, on peut constituer un arbre de parenté phénoménologique, pas un arbre de descendance ; or, et c'est une surprise qui a évidemment ravi les premiers expérimentateurs, on découvre que cet arbre a la même forme que l'arbre découvert par les paléontologues. C'est extrêmement frappant, puisque dans un cas, le cas de l'analyse des gènes par la biologie moléculaire, les objets étudiés sont à la fois synchroniques et moléculaires alors que, dans l'autre cas, dans le cas de la paléontologie, ils sont diachroniques et macroscopiques. Si l'on peut parler de preuve, c'est bien là une preuve de l'idée d'évolution des espèces, mais c'est aussi un moyen de dater les objets. Je ne vais pas discuter ici du problème de l'horloge moléculaire, mais il est possible, en première approximation, et c'est là aussi une coïncidence assez intéressante, de se rendre compte que tout se passe comme si les différents êtres vivants évoluaient grossièrement à la même vitesse. C'est très surprenant, puisque cela veut dire qu'une bactérie, qui peut produire dix mille générations par an varie à la même vitesse que l'Homme qui se reproduit tous les vingt ou vingt-cinq ans. L'une des raisons de ce paradoxe est probablement que l'une des bases des acides nucléiques est sensible à des paramètres physiques, probablement aux rayons cosmiques qui inondent la Terre en permanence. Sous leur action, la cytosine est désaminée. Cela conduit systématiquement à certains types d'erreurs. Dans ce cas, ce qui compte, c'est l'aptitude à les corriger ; or, cela se fait pour l'essentiel au moment de la reproduction. Par conséquent, si l'être considéré se reproduit souvent, il a de meilleures chances de corriger ses erreurs et, en effet, on observe que les bactéries font plutôt moins d'erreurs (évoluent moins vite), bien qu'elles aient beaucoup plus de générations, que les êtres humains par exemple. Il y a bien sûr une énorme controverse autour de l'horloge moléculaire parce qu'il y a des gènes qui évoluent plus vite que d'autres, etc., mais en gros, elle peut donner une grossière image de ce qu'est l'évolution.
Je termine à présent. Mais je voudrais vous mettre en garde sur un point encore : j'ai mentionné la constitution d'arbres, la reconstitution d'un arbre généalogique à partir de la comparaison de génomes entre eux. Cette comparaison repose sur la mesure des distances entre objets et sur l'hypothèse implicite de la dichotomie, c'est-à-dire qu'on suppose que chaque filiation se fait en un point en donnant deux descendants différents. Or, — c'est un point très important et je crois d'ailleurs que ceux qui s'opposent à propos de filiations différentes, s'ils en avaient conscience, arriveraient à concilier leurs points de vue — je pense que cette hypothèse est une hypothèse inadéquate. Et je vais vous dire pourquoi : quand on commence à étudier les systèmes vivants et, en particulier, leurs protéines, on se rend compte d'une particularité tout-à-fait originale à savoir leur robustesse qui se manifeste en ce qu'elles peuvent comporter un nombre d'erreurs absolument incroyable avant de perdre leur fonction. C'est une observation assez surprenante qui laisse penser qu'on va avoir de nombreuses règles à découvrir dans la genèse des protéines elles-mêmes, dans leur organisation. Or, une idée répandue, que j'ai d'ailleurs contribué à répandre, est celle de l'avalanche d'erreurs : c'est l'idée que si la machinerie de synthèse des protéines se trompe, alors, puisque les protéines reproduisent l'ADN qui les spécifie, les erreurs vont s'accumuler et cela va mener à la catastrophe. C'est pourtant très visiblement inadéquat. En effet, le système peut continuer à subsister malgré un grand nombre d'erreurs. Il existe un mutant qui ne sait pas corriger les erreurs de réplication, c'est-à-dire que la copie de l'ADN n'est pas fidèle. Normalement, il existe un système de relecture qui permet de corriger les erreurs, et ce système est absent dans ce mutant. Or, ce mutant 1) est viable, ce qui est frappant, mais 2), et c'est à mon avis la chose qui, de très loin, est la plus frappante, il n'engendre pas deux individus identiques. Par conséquent, si ce genre d'événement arrive, il donne naissance nécessairement à toute une série de descendants très différents les uns des autres. On peut parfaitement imaginer qu'au cours de l'évolution, par exemple, avant l'origine de l'Homme et du chimpanzé — il existe une querelle qui agite les chercheurs dans ce domaine, pour savoir qui est antérieur du chimpanzé ou du gorille — on peut penser que, et il y a beaucoup de circonstances où existe une situation de ce genre, la population issue d'un même ancêtre est si variée que deux individus ont des génomes extrêmement distincts les uns des autres. C'est donc évidemment une situation instable, on le comprend très bien. Mais si cela arrive, cela ne peut pas durer très longtemps à l'échelle des temps géologiques. Et cela peut donner plusieurs branchements provenant approximativement au même moment d'un même ancêtre. Pour terminer ce propos, je rappelle aussi que lorsqu'on veut remonter à l'origine, on peut bien sûr faire des comparaisons, mais il faut être très conscient des contraintes qu'on s'impose dans la reconstitution des arbres et la reconstitution des hypothèses. La dichotomie en est une comme l'est cette hypothèse que j'ai mentionnée tout au début, la relève génétique : bien sûr, il peut exister des choses qui ont complètement effacé leur passé et, dans ce cas, nous n'en aurons jamais la mémoire.
Je voudrais remercier Monsieur Danchin pour cette conférence extrêmement brillante et passionnée, quelquefois un peu difficile à suivre, mais même si les exemples n'étaient pas toujours familiers à tout l'auditoire, les conclusions que vous en tiriez étaient, elles, beaucoup plus claires. Alors, je suppose qu'il va y avoir des questions.
Monsieur Danchin, je voudrais vous poser deux questions. Comment peut-on expliquer le choix des acides aminés lévogyres dans les protéines ?
A ce sujet, je crois que les choses sont en fait assez simples. On a souvent tendance à penser que ce qui est stable est la symétrie. En fait, dans un très grand nombre de cas, ce n'est pas vrai. Ce qui peut être stable, c'est le fait de ne pas être symétrique. Si l'on a initialement un ensemble de molécules chargées posées sur une surface, par exemple deux isomères en symétrie miroir, comme le sont la main gauche et la main droite, elles peuvent se concentrer localement à un moment donné en tirant à pile ou face, vous pouvez l'observer sans aucune difficulté. Ce sera par exemple l'un des deux isomères, le droit ou le gauche. Et si cet objet a commencé localement à amorcer un certain nombre de synthèses, sa propre activité lui permettra de sélectionner les objets de sa propre famille. C'est typiquement l'exemple que j'ai donné tout-à-l'heure de la route et de la conduite à gauche ou à droite : on conduit d'un côté ou de l'autre, mais d'un seul. Si l'on a fait un choix, on est piégé par ce choix. Je crois que ce choix est contingent, c'est-à-dire qu'on pourrait imaginer un autre monde où tout soit symétrique de ce qui est dans le nôtre. Et d'ailleurs, puisque vous me posez cette question, elle conduit à une autre question évidente : est-ce qu'il y a de la vie ailleurs ? Posée de cette manière, la question devient très riche puisque, dans les prémisses de ce que je vous ai dit, on voit très bien que tout ce qui a conduit à la vie peut commencer assez facilement et même avec une probabilité très élevée. Et donc, on peut parfaitement imaginer — et je pense qu'on doit l'imaginer — qu'il y a une chimie du carbone dans un grand nombre d'environnements célestes, et qu'elle produit des molécules un peu dans le genre de celle que j'ai imaginées au début. Mais le problème devient tel que, à chaque fois qu'il se produit quelque chose de nouveau, cela crée une contrainte originale, c'est-à-dire que l'évolution du système avance selon les branches d'un arbre, sans pouvoir revenir en arrière. Et je ne crois pas qu'on puisse savoir calculer avec vraisemblance la probabilité d'un retour en arrière une fois qu'ont été utilisées un certain nombre de branches. En ce qui concerne l'origine de la vie, il nous faudrait connaître le nombre des branchements possibles d'une chimie du carbone initiale. Or, on n'en a aucune idée, et par conséquent, bien qu'on puisse dire qu'il y a certainement ailleurs une chimie du carbone évoluée, il ne nous est pas possible de savoir si cette chimie est susceptible d'avoir évolué assez loin pour séparer mémoire et fonction, ce qui est finalement, comme je vous l'ai dit, le cœur du problème. Et je dirais même que la question se double pour nous d'une deuxième question : l'origine de la vie est une question, mais l'origine de l'Homme en est une autre, indépendante, puisque, je vous l'ai dit, il a été inventé un deuxième code, celui du langage. Je crois d'ailleurs que si, sur terre, on faisait disparaître aujourd'hui l'ensemble des mammifères, ce qui n'est pas impossible, il n'est pas sûr du tout que ces mammifères pourraient apparaître à nouveau : une fois qu'on a avancé — c'est essentiel dans l'idée arborescente des choses — une fois qu'on a avancé le long des branches, on ne peut pas revenir en arrière, et par conséquent, on ne peut pas faire de calcul du type calcul de probabilités. Le fait qu'il y ait un choix, est un fait qu'on peut considérer comme contingent a priori et nécessaire a posteriori.
Monsieur, vous avez parlé au point de vue des idées reçues, vous avez dit que vous étiez contre et je voudrais vous demander dans ces idées reçues, il y a celle du fait que s'il n'y a plus de molécules dans une dilution, il ne peut plus y avoir d'action.
Ce n'est pas très difficile. La première chose que je voudrais vous dire, c'est que je n'ai pas dit que j'étais contre les idées reçues, ce serait absurde. Nécessairement, je n'ai initialement que des idées reçues. Ce que je dis, c'est qu'il est nécessaire de savoir revenir sur les idées reçues. Et c'est pour cette raison que je suis revenu sur l'épigraphe du livre de Jacques Monod. Ce qui, à ma connaissance, n'a été fait par personne bien qu'il ait été lu par des centaines de milliers de personnes. Cela en dit long d'ailleurs sur la façon dont nous lisons les livres. En préalable à la question que vous me posez, je vous rappelle qu'on peut toujours inventer à peu près n'importe quoi. Dans le cas précis que vous mentionnez, il est tout-à-fait clair que les expériences, telles qu'elles ont été construites, n'auraient pas été admises chez le moindre étudiant, tellement elles sont dépourvues de contrôle (7). En bref, il n'y a pas d'eau pure. Et il aurait fallu pouvoir en contrôler d'abord la pureté avant d'aller plus loin. C'est là essentiellement ma critique sans même contester le modèle expérimental, qui est lui-même contestable. Je vais vous donner un exemple très simple, qui met en jeu notre propre discipline. Cet exemple est très amusant, c'est la découverte du rôle d'un ion, l'ion fluorure. On voit encore dans des publications aujourd'hui, bien qu'il y ait près de dix ans qu'on en a découvert la cause, je crois que c'est Marc Chabre à Grenoble qui a fait cette découverte — on voit, dans un très grand nombre d'analyses biochimiques, l'étude d'un effet du fluorure. Et l'on parle de l'effet fluorure, c'est bien classé, inventorié, référencé. Vous savez que, dans les laboratoires, on utilise nécessairement des objets qui nous sont fournis par des fournisseurs variés et qu'il y a là des effets de mode ou des progrès. On est passé un jour, c'est de plus en plus fréquent maintenant, du verre au plastique. Et quelqu'un, dans un laboratoire, faisait des expériences avec l'ion fluorure dans de l'eau aussi propre que possible. Mais, et c'est là un des problèmes majeurs, il n'y a pas, le plus souvent, d'eau qui puisse avoir un degré de pureté comparable avec ce qui est demandé lorsqu'on veut y faire de grandes dilutions. L'expérience avec le fluorure a donc été faite mais, ô surprise, sans résultat. C'était pourtant une expérience qui avait été reproduite en laboratoire pendant des années. Ce qui avait changé, c'était le matériel dans lequel on stockait l'eau. Ce matériel était du plastique au lieu d'être du verre. Et l'on s'est rendu compte qu'en fait, quand on met du fluorure dans l'eau, si cette eau est conservée dans du verre, le fluorure captera les ions aluminium de la surface ce qui fabrique les ions AlF4–. Or, l'ion AlF4– a une architecture qui ressemble au phosphate et il se trouve donc que l'effet fluorure qu'on décrivait depuis des années est en fait un effet phosphate. Tout cela pour dire que, tant qu'on n'a pas fait le minimum de contrôles de ce type où la pureté est un critère essentiel, on devrait s'abstenir de publier. Je dois vous dire que la personne que vous mentionnez à propos de ces expériences a publié, bien avant que ne sortent ces résultats invraisemblables, des expériences dans lesquelles l'eau qui était utilisée était l'eau du robinet. A mon sens, cela ne mérite pas plus de commentaires. Donc, ce n'est pas une question d'idées reçues, c'est simplement une question de propreté des expériences. Il est inutile de s'interroger sur les conclusions d'expériences mal conduites. Il faut d'abord bien les faire.
Est-ce qu'on peut dire [inaudible] s'éclater, si j'ose dire et avoir [inaudible]. actions... ?
Si, le métabolisme, c'est cela. C'est la décomposition des molécules en éléments qui vous donne l'eau. Métaboliser une molécule, c'est la faire passer d'une espèce moléculaire à une autre espèce. C'est exactement ce que fait le système vivant, c'est clair.
Vous avez donné comme définition de la vie la séparation de la fonction et de la mémoire.
Enfin, une des définitions...
Est-ce que c'est une définition arbitraire, ou est-ce que vous avez élaboré là-dessus, pourtant vous mettez l'accent là-dessus ?
Vous me posez là une question qui pose un problème énorme. Et je crains d'avoir à répondre pendant très longtemps. Un système vivant, c'est essentiellement une machine, qui construit avec des objets élémentaires, en petit nombre, d'autres objets qui ont la propriété de pouvoir prendre une grande variété de formes, un peu comme avec quelques boulons et quelques parpaings, on peut faire des constructions absolument extraordinaires. Toute la question qui se pose est donc, si l'on a une machine de ce type, est-ce qu'on peut trouver un enchaînement d'ordres — je prends volontairement une image informatique — un enchaînement d'ordres qui permet de mettre les objets bout à bout pour construire de cette manière des édifices compliqués. L'idée de la mémoire est alors l'idée du programme, de la suite des ordres permettant cette construction. On peut considérer un système vivant comme un automate, un automate qui a la propriété originale d'avoir séparé la mémoire permettant de spécifier l'enchaînement des instructions de l'objet que l'on manipule et qui est créé par ces instructions (mémoire et fonction). On s'aperçoit que cet ensemble possède des propriétés émergentes, c'est-à-dire que le fait de pouvoir construire des objets avec la possibilité d'y introduire des erreurs, des variations permet de créer des objets nouveaux sans commune mesure qualitativement avec les objets qui ont précédé. Je ne sais pas comment vous donner une autre analogie. Prenons simplement deux transistors, vous les connectez n'importe comment, cela ne donne rien d'intéressant. Mais si vous les connectez d'une certaine manière avec les deux ou trois connections appropriées et vous fabriquez un amplificateur, c'est-à-dire un système irréductible aux précédents. Pour en revenir à l'idée de codage, c'est ce qui permet au système de revenir sur lui-même. Il se boucle puisque l'ensemble des instructions spécifie les objets qui manipulent l'ensemble qui, à son tour, crée les instructions. Et c'est essentiellement cela qui fait que le système qui possède ces propriétés de codage produit sans cesse des propriétés émergentes, c'est-à-dire irréductibles à celles qui lui ont donné naissance. Et qui plus est, nous, êtres humains, employons un deuxième code, le langage. Et cela crée une situation du même ordre, c'est-à-dire que nous créons des objets qui sont réellement, profondément, des productions émergentes, des artifices. Je ne sais pas, il faudrait le demander aux physiciens, mais il ne me semble pas qu'un faisceau laser, par exemple, soit quelque chose qui existe dans la nature. Typiquement, c'est le produit d'une réflexion et, pourtant, c'est un objet qui a une action, qui a un certain nombre de propriétés engendrées par le fait qu'on a pu produire un retour sur soi. Un autre exemple : si vous prenez — c'est l'exemple que donne Douglas Hofstadter, je pense qu'il n'est pas mauvais — une caméra vidéo et un écran de télévision. Vous prenez la caméra, vous regardez et vous voyez l'image à l'écran, puis vous faites la chose suivante, vous orientez la caméra de façon à ce qu'elle regarde l'écran. Alors, vous créez des formes nouvelles, inattendues : spirales, figures oscillantes etc.. Ainsi, le fait de créer une boucle de ce genre, fait sortir le système de lui- même et engendre du neuf. Et c'est ce qui me paraît tout-à-fait particulier au système vivant. Je ne crois pas qu'on en ait du tout exploré les conséquences dans la production des formes et des dynamiques. C'est-à-dire que ce n'est pas un paradoxe de la matière, c'est une de ses propriétés.
Mais je ne peux pas du tout faire la comparaison ainsi que vous le demandez. Si vous voulez, les modèles astrophysiques locaux, du moins ce que j'en connais, sont des modèles qui sont essentiellement tournés vers l'extérieur et sans frontières. Ce dont je vous parle a une particularité, c'est que l'ensemble des objets pertinents réuni à l'intérieur d'une membrane forme un individu. C'est évidemment tout-à-fait essentiel.
Vous semblez douter du programme de recherches sur le génome, sur deux aspects. Le premier, c'est que déterminer la séquence à raison de 6 millions de paires de bases à raison de cinquante mille paires par personne et par an est considérable, et deuxièmement quel serait le but d'un programme de recherches ?
Vous me posez là une question qui a des implications politiques, ce qui me met relativement mal à l'aise puisque je ne possède pas tous les éléments pour vous répondre. Ce que je peux vous dire, c'est que, telle que Watson l'a lancée aux Etats-Unis, l'idée était un peu d'accomplir un exploit. Il s'agissait d'un grand projet comme aller sur la Lune, et l'idée n'était pas fondamentalement réfléchie du point de vue de la biologie. Maintenant, il est évident que connaître mieux notre génome a un intérêt. Ce que je mettais en doute, mais ce n'est d'ailleurs pas ce qui se fera dans un avenir proche, n'était pas l'étude de notre génome mais plutôt l'idée de commencer par en faire la détermination de la séquence. Je ne crois pas que ce soit véritablement un problème essentiel dans l'immédiat. En revanche, déterminer quels sont les gènes importants, ceux qui sont par exemple utilisés dans telle ou telle classe de cellules, faire le tri des ADN complémentaires, voilà ce qui serait un projet qui me paraîtrait tout-à-fait raisonnable. Un autre projet, en cours, qui n'est pas absurde, mais qui à mon avis, pose des questions éthiques redoutables, est la détermination de la cartographie du génome humain. C'est évidemment un problème intéressant mais pour lequel, à mon avis, on est obligé de se soucier des points de vues éthiques. C'est pour cette raison que je pense que le projet de séquençage du génome humain a deux handicaps. Il y a 1. l'inadéquation de la détermination de la séquence totale en tant que projet scientifique et 2. les problèmes éthiques soulevés méritent quand même une discussion et une réflexion très profondes. Ce qui ne veut pas dire qu'il ne faut pas lancer un programme de recherches dans ce domaine.
Vous rejetez l'expérience qui a été faite concernant les acides aminés à partir de la soupe primitive. Est-ce qu'il faut les rejeter totalement ou est-ce qu'une combinaison des théories sur les pierres et le métabolisme de surface et les expériences qui ont été faites permettent un lien entre les deux ?
Si, il est tout-à-fait possible de les relier. La question telle qu'elle était posée, c'est-à-dire s'imaginer être Dieu le Père et créer le système vivant me paraît une question inadéquate. C'est essentiellement cela que je refuse dans l'idée de la genèse prébiotique. A l'inverse, je pense qu'il vaut mieux partir de ce que nous connaissons, essayer de remonter et de voir où l'on doit s'arrêter. S'il y a relève génétique, comme je vous l'ai dit, il y a un endroit où l'on sera forcément arrêté, quel que soit notre imaginaire pour la construction des soupes. Je n'ai pas envie de rejeter radicalement les modèles anciens, je pense simplement qu'ils sont essentiellement non pertinents.
Cela peut être pris comme partiel.
Cela peut être pris comme partiel, mais je ne vois pas comment cela peut apporter quelque chose de constructif à la question. Sauf peut-être une chose, c'est que cela démontre que la chimie du carbone est facile. Qu'il n'y a pas de paradoxe dans la chimie du vivant.
Vous avez parlé de la dissemblance dans les génomes et il y a le cas du génome des jumeaux semblables.
Oui, les génomes homozygotes sont effectivement semblables entre eux, mais ils possèdent des chromosomes du père et de la mère. Par conséquent, si l'on prend un morceau du gène de l'un, du père ou de la mère, on rencontrera des problèmes d'identification parce qu'il y aura 5 % de polymorphisme. Une façon de procéder, qui d'ailleurs est utilisée, c'est d'utiliser la consanguinité "naturelle" comme les cas incestes de père en fille, après plusieurs générations lorsque cela existe. Cela se fait car cela conduit souvent à l'apparition de maladies génétiques.
Moi, j'ai envie de vous poser une question qui sera moins scabreuse. C'est quelque chose qui m'a beaucoup intéressé dans votre méthode. C'est lorsque vous évoquez la façon dont certaines synthèses se font et que vous dites que pour un chimiste d'aujourd'hui, elles sont absurdes. Et cela me rappelle ce qui a été dit depuis longtemps, à savoir qu'il y a dans les organismes vivants, un certain nombre de structures qui choqueraient un élève de première année des Arts et Métiers. Alors, une des explications traditionnelles, surtout mise en avant par les paléontologistes, c'est l'histoire des organes vestigiels et des contraintes que l'existence de certains ancêtres a posées sur le développement d'organismes ultérieurs. J'ai eu le sentiment que vous utilisez au fond un type de raisonnement analogue.
C'est vrai. Je n'ai pas mentionné une part extrêmement importante de l'évolution, qui explique probablement en grande partie, ou qui rend compte en grande partie de la robustesse des systèmes vivants. C'est ce qu'on pourrait appeler l'opportunisme moléculaire. Je vais essayer de vous l'expliquer. S'il y avait brutalement un coup de vent dans cette pièce, mes notes commenceraient à s'envoler, et j'utiliserais immédiatement ce paquet (c'est-à-dire des crayons) dans une fonction qui n'a rien à voir avec leur destination comme presse-papier. C'est typiquement quelque chose qui arrive tout le temps. C'est un point extrêmement important qui explique effectivement la bizarrerie qu'un ingénieur des Arts et Métiers remarquerait, la bizarrerie de la construction d'un certain nombre d'objets biologiques. Le fait d'avoir à sa disposition un certain objet qui a une fonction, mais qui peut en acquérir une autre pour une raison purement contingente, mais adaptée aux circonstances, peut être utilisé ensuite, servir de point de départ pour une évolution. Je vais vous donner un exemple qui est très frappant, et à mon avis tout-à-fait merveilleux, à savoir le cas du cristallin de l'œil. Vous savez que les yeux des vertébrés, mais aussi les yeux de certains invertébrés ont un cristallin. Et l'une des particularités de ce cristallin est d'être rempli de telle manière que ses protéines sont assemblées de façon vitreuse, ce qui lui donne sa transparence. Je vous ferai remarquer d'abord qu'en ce qui concerne la possibilité pour des protéines, pour un concentré de protéines, d'avoir une apparence vitreuse, il n'y a pas de paradoxe: vous connaissez très bien le cas du blanc d'œuf : un blanc d'œuf est un ensemble de protéines très concentré et est effectivement très transparent jusqu'à ce qu'on le dénature en le chauffant. Bien. Dans le cas du cristallin, on pourrait se demander si, depuis l'origine des vertébrés, nous avons tous véritablement hérité d'un même ensemble qui a été choisi pour ses propriétés optiques. Or, il y a quelques années, un ensemble de chercheurs a étudié les protéines qu'on appelle cristallines, des cristallins de la grenouille, de l'Homme, de l'oiseau, etc. Alors, à leur grande surprise, ils ont découvert que ces protéines, lorsqu'ils les étudiaient, étaient chez un animal, une enzyme ordinaire, c'est-à-dire une protéine douée d'une activité enzymatique du métabolisme n'ayant rien à voir a priori avec un cristallin, chez un autre animal, une autre enzyme, et chez un troisième animal, encore une autre enzyme, comme si le système avait été inventé un grand nombre de fois et que, en réalité, faire un cristallin, était quelque chose d'assez facile ; il suffisait d'y mettre beaucoup d'une protéine quelconque. Ils ont découvert ensuite une deuxième chose assez remarquable, et qui va me ramener à la question du génome humain, c'est la chose suivante : vous voyez bien que le problème des cristallins n'est pas seulement d'être transparents mais aussi de garder leur transparence au cours d'une durée de vie longue. Chacun sait que la cataracte est un accident fréquent lié à l'âge. Cela montre bien qu'il y a une instabilité dans la structure des protéines qui fait, qu'au bout d'un certain temps, ces protéines se dénaturent. Mais ce qui vient d'être découvert à ce propos, c'est quelque chose de tout-à-fait récent et de tout-à-fait merveilleux. On sait qu'il existe deux grandes classes de protéines, très probablement un peu plus. Certaines sont des protéines qui contiennent peut-être en elles-mêmes toute leur aptitude à prendre leur architecture, je m'explique : ce sont des suites d'acides aminés qui reprennent leur forme lorsqu'on les déroule et qu'on les remet dans l'eau. Ce sont les protéines les plus classiques. On a découvert récemment que ce n'est pas vrai de toutes les protéines et qu'au moins la moitié d'entre elles (peut-être toutes en réalité) ont besoin en fait d'échafaudages — appelés "chaperons" parce qu'elles accompagnent les protéines — pour pouvoir se replier correctement. Alors, ce qui est intéressant, c'est que ces protéines-échafaudages, qu'on découvre tous les jours : il y en a maintenant un très grand nombre, ont un rôle absolument fondamental. Ces protéines-échafaudages sont souvent liées à des conditions de stress pour les cellules, c'est-à-dire quand on monte brutalement la température, qu'on les expose à l'alcool, ou qu'on les expose à trop d'oxygène, des situations de ce genre qui sont évidemment dénaturantes. Il existe ainsi un dispositif de protection qu'on appelle les protéines de choc thermique. Parmi ces protéines, certaines ont une activité enzymatique, par exemple des activités essentielles et d'autres sont des échafaudages. Or, on a découvert que ce qui remplit les cristallins est justement presque toujours un ensemble de protéines de choc thermique, c'est-à-dire en fait, des enzymes plus les chaperons qui leur correspondent. Et les chaperons sont là précisément permettant de renaturer les protéines quand elles se dénaturent, ce qui explique leur très longue stabilité... Et là, je reviens au génome humain. Vous avez remarqué que cette évolution qui a donné naissance au cristallin est une évolution par perte d'une fonction. Normalement, les protéines de choc ne sont induites qu'en présence d'un choc. Sinon, elles sont absentes. Les gènes sont silencieux. Donc, dans ce cas-là, ce n'est évidemment pas ce qui se produit puisqu'elles sont là en permanence, puisqu'elles permettent de construire un cristallin. C'est donc la perte de la fonction contrôle, de l'expression des gènes correspondants qui a permis de créer le cristallin. Et c'est pour moi une morale extrêmement importante : même la perte d'un gène peut être bénéfique.
J'imagine que deux individus ne sont jamais identiques parce qu'ils n'ont jamais la même chaîne d'ADN, parce que trois milliards de caractères de combinaisons, effectivement cela donne une idée des chances extrêmement improbables d'avoir deux individus identiques. Par contre, on peut avec des probabilités beaucoup plus grandes trouver des morceaux d'ADN communs. Alors est-ce que ceci n'expliquerait pas des phénomènes qui sont hors du domaine de la science comme ceux des personnes qui ont le sentiment d'avoir vécu une expérience communune, est-ce qu'ils n'ont pas un morceau de chaîne qui correspond à la mémoire de la vie passée ?
Oui, je vois ce que vous voulez dire. Je vais vous surprendre. En réalité, la différence, l'identité est quelque chose de bien plus extraordinaire encore que ce que vous pensez : deux bactéries ayant le même génome strictement, et plongées dans le même environnement ne sont pas identiques. Et je vais vous dire pourquoi. Il n'y a jamais identité, dans aucun cas. C'est un point essentiel, même si les génomes sont strictement identiques, les environnements strictement identiques. La raison en est très simple. On a tendance en général à traiter, à tort, les êtres vivants comme relevant des lois les plus habituelles de la physique ; or, les lois habituelles de la physique sont souvent liées à des phénomènes macroscopiques et statistiques. Or, précisément dans les systèmes vivants, ce devrait être interdit. En effet, le nombre des objets considérés, la plupart du temps est très petit. Comme il est très petit, il ne relève pas de la statistique des grands nombres, du moins pas de façon usuelle. Cela veut dire la chose suivante, si vous avez un gène de contrôle par exemple, comme dans le cas de l'assimilation d'un sucre, le lactose, la protéine qu'il spécifie existe en très petit nombre d'exemplaires dans la cellule, une dizaine de molécules au plus. Cela signifie que le messager correspondant est présent en un unique exemplaire. Les variations d'un individu à l'autre seront donc du type présence ou absence, c'est-à-dire donnant des phénotypes très différents, et ces variations seront sensibles à d'infimes perturbations du milieu. Ainsi, comme cela se reproduit très souvent, deux individus ne seront jamais identiques. Ainsi, il ne peut pas y avoir de mémoire génétique du passé autre que très grossière : mais il est clair que des classes de mémoire existent, ne serait-ce que dans la reconnaissance du bon partenaire sexuel pour un individu donné. Mais ce n'est jamais la "vie antérieure".
![]()