Santiago RAMON Y CAJAL
L'inné et l'acquis : une théorie sélective de l'apprentissage
Antoine Danchin
Cet article a été publié initialement dans le magazine La Recherche en février 1974. Il fait suite aux travaux d'un groupe animé par Jean-Pierre Changeux, Philippe Courrège et moi-même, qui se réunissait tous les mercredis après-midi à l'Institut de Biologie Physico-Chimique pour formaliser les propriétés sélectives de l'évolution des réseaux de neurones, et qui se déroulait en parallèle avec mon activité de coordinateur au Centre Royaumont pour une Science de l'Homme. Il reprend tout son intérêt au moment où il devient possible de comprendre comment la catégorie du Réel que nous appelons "information" s'articule avec les catégories standard, masse, énergie, espace et temps. Avec notre vision actuelle du démon de Maxwell, l'apprentissage se produit à la suite d'un mécanisme sélectif qui doit utiliser de l'énergie pour éviter de faire disparaître les seules synapses qui fonctionnent assez souvent. Le mécanisme correspondant, codé par des gènes, doit être identifiable, de même que la source d'énergie qui produit cet évitement de la dégradation.
Après plus d'un siècle de recherches en neurobiologie, il apparaît désormais évident que les cellules nerveuses (les neurones) et plus particulièrement les contacts que ces cellules forment entre elles, ou synapses, ont une fonction primordiale dans le traitement de l'information qui, sous forme d'impulsions électrochimiques, parcourt le réseau nerveux.
Dès 1895, dans son Esquisse d'une psychologie scientifique, Freud essaie d'expliquer la mémoire avec les connaissances biologiques de son temps. Il constate que les neurones doivent être impressionnés sans pourtant être altérés, ce qui le conduit à refuser la distinction entre des neurones de la perception et des neurones du souvenir. Il imagine alors que c'est le chemin tracé au travers des zones de contact préexistantes (que nous appelons les synapses) qui va permettre la persistance de la mémoire : celle-ci est représentée par la différence entre l'ensemble des chemins fournis par l'hérédité à l'individu et les chemins effectivement acquis au cours de l'expérience individuelle; et pour Freud le moteur de cette sélection est la répétition, qui permet de frayer la voie qui est la mémoire.
Depuis, de nombreuses observations ont permis de préciser ces idées.
Une nouvelle théorie de l'apprentissage dans les réseaux nerveux
Pour décrire les capacités du système nerveux individuel, il est commode de distinguer trois niveaux : la structure du système, c'est-à-dire son organisation spatiale, avec les différents types de cellules et de connexions; son fonctionnement, c'est-à-dire les règles qui définissent la production et l'intégration de l'activité électrochimique du réseau nerveux, et enfin son comportement, c'est-à-dire la façon dont un individu contrôle, à l'aide de son système nerveux, son interaction avec son environnement.
L'observation générale du système nerveux central, et de ses propriétés, montre, à ces trois niveaux, un programme génétique particulièrement rigide (voir l'article de Jean-Pierre Changeux, 2); la rigidité de la structure est bien mise en évidence par l'existence de mutants qui sont des variants héréditaires d'un type anatomique originel (au niveau de l'organisation globale, d'une classe de neurones, d'une classe de synapses...). De même les règles qui prescrivent la spécificité de la connectivité sont établies avec une grand précision, comme les règles d'apparition des potentiels d'action (influx nerveux) et leur rythme d'activité en fonction du développement de l'individu. Enfin, par voie de conséquence, le comportement lui-même est dans une très grande mesure génétiquement déterminé.
Cependant, si en général pour les petits invertébrés la rigidité du programme génétique apparaît absolue, au fur et à mesure que le système nerveux se complique, une certaine fluctuation apparaît : ce n'est qu'un schéma général de l'organisation des trois niveaux mentionnés qui est fourni par le programme génétique. Et l'évolution des formes de l'apprentissage vers les formes les plus complexes semble aller de pair avec l'apparition de cette fluctuation. On peut noter en outre deux caractéristiques de cette fluctuation : d'une part, elle apparaît surtout au début de l'ontogenèse, durant une période critique, mais persiste parfois chez l'adulte; d'autre part, la stabilité de la structure semble liée au fonctionnement général du système : les expériences de déprivation sensorielle (où l'on prive un jeune animal d'une interaction avec son environnement) montrent très souvent une dégénérescence profonde de l'organisation du réseau nerveux.
Ces observations sont à la base d'une théorie de l'apprentissage dans les réseaux nerveux dont les fondements viennent d'être publiés par J.P. Changeux, P. Courrège, A Danchin. C'est une théorie de l'épigenèse des réseaux nerveux.
L'épigenèse est, au cours du développement de l'être vivant, la formation d'une organisation par interaction avec l'environnement. L'apprentissage est considéré comme l'acquisition, au cours d'une interaction avec l'environnement, d'une propriété associative bien définie et stable, et discutée en termes d'épigenèse. La mémoire est alors un apprentissage susceptible d'être rappelé (il s'agit ici de mémoire de longue durée).
Le modèle est construit suivant la méthode axiomatique, c'est-à-dire que la définition et l'étude du modèle mathématique sont soigneusement distingués de sa signification biologique (1).
Ainsi la rigueur et la cohérence interne du modèle mathématique ne peuvent être affectés par la pensée biologique tandis que les problèmes posés par la biologie ne peuvent être déformés par la rigidité ou la schématisation grossière du modèle. Ce sont des énoncés particuliers, appelés interprétations, qui permettent de coupler la biologie à la formalisation mathématique. En ce sens, cette méthode, qui est fondée sur des postulats biologiques qui deviennent les interprétations d'axiomes mathématiques, est totalement opposée à la méthode usuelle qui consiste à exhiber un objet mathématique dont on décrit les propriétés en termes pseudo-biologiques pour ensuite assimiler, par un raisonnement analogique, le modèle et la réalité expérimentale.
Postulats biologiques et axiomes mathématiques
Les postulats biologiques qui nous semblent pertinents, et suffisants pour servir de base aux caractéristiques fondamentales de l'apprentissage, de la mémoire, et sans doute des propriétés les plus évoluées du système nerveux central des vertébrés supérieurs, sont les suivants :
I. Ce sont les neurones qui sont responsables du traitement de l'information accompli par le système nerveux : il n'est retenu que le caractère discontinu des influx qu'ils créent, transmettent, intègrent et propagent.
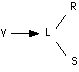
II. Ce sont les contacts entre les neurones ou synapses qui sont les intermédiaires de tous les transferts d'information à travers le système. Ces synapses peuvent exister en trois états au moins (plasticité synaptique) : labile (L), stable (S) et dégénéré (R); seuls L et S transmettent l'influx nerveux, et les transitions permises sont :
III. L'évolution de l'état de chaque synapse est réglé par l'ensemble des influx qui arrivent sur le neurone post-synaptique, pendant un intervalle de temps de longueur déterminée (on dit que le soma neuronique a un pouvoir évoluteur).
IV. La connectivité maximale ainsi que les étapes du développement du réseau, de même que le pouvoir évoluteur et intégrateur de chaque soma neuronique, sont déterminés de façon rigoureuse par le programme génétique : cela constitue l'enveloppe génétique du réseau.
V. L'apprentissage est considéré comme un aptitude acquise associée à la variabilité de l'organisation des connexions du système. Ce qui est inné dans l'enveloppe génétique.
Ce qui est inné est donc l'enveloppe génétique et ce qui est acquis est une propriété associative stable (voir aussi (2)). L'apprentissage est le processus par lequel une certaine classe d'interactions avec l'environnement conduit à la mise en place de cette propriété associative. D'après la théorie proposée, celle-ci se trouve inscrite dans la structure (et le fonctionnement) du réseau nerveux en une forme particulière de l'organisation du réseau. Cette forme résulte en général de la stabilisation par le fonctionnement de certains chemins stables parmi le très grand nombre de possibilités fournies par l'apparition des synapses labiles au cours du développement. D'une façon imagée, on peut représenter le réseau nerveux comme un immense filet à trois dimensions dont les nœuds seraient connectés les uns avec les autres en très grand nombre : au cours de l'interaction avec l'environnement, une partie des connexions se trouvera coupée, et ainsi une image particulière de l'environnement est sculptée dans la masse du réseau.
Les réalisations d'un programme neuronique
Du point de vue formel, on étudie un objet mathématique appelé un programme neuronique R = (C, Σ ,Θ, n, Φ, Δ) où (C, Σ) est un graphe particulier, où l'on peut distinguer des entrées et des sorties, que l'on interprète comme la structure connexionnelle maximale possible; Θ représente pour chaque connexion le délai de transmission de l'influx nerveux entre son point d'émergence et le neurone suivant; Φ représente le pouvoir d'intégration qui permet à chaque neurone d'envoyer un influx nerveux en fonction du message total qu'il reçoit pendant la durée n; Δ, enfin, représente le pouvoir d'évolution de chaque neurone sur chacune de ses synapses afférentes, en fonction du programme génétique (la croissance est prise en compte en supposant qu'une connexion est dans un état néant N avant d'apparaître dans un état labile) et du message total afférent pendant la durée n.
L'objet de la théorie est d'étudier les propriétés des réalisations de ce programme neuronique en fonction d'un multi-message qui est imposé sur les entrées du graphe neuronique et qui représente la pahse d'apparence "instructive" de l'apprentissage. En particulier, une classe de programmes neuroniques est susceptible d'acquérir une T-T' compétence, c'est-à-dire qu'au bout d'un temps fini, des messages d'un type T donné sont transformés en des messages d'un type T' stable à la sortie : en général, une telle compétence ne reste stable qu'à la condition qu'un fonctionnement minimal soit imposé à l'entrée du réseau; d'ailleurs on observe que, dans la période d'acquisition de la T-T' compétence, un faible fonctionnement conduit à une très large dégénérescence (cela correspond bien aux expériences de déprivation sensorielle). Ainsi, l'expression du programme génétique ne peut dans certains cas se développer correctement qu'à condition que le réseau nerveux fonctionne. C'est là qu'intervient l'environnement de façon privilégiée, et c'est là que va se marquer l'empreinte qui sera l'acquis. Cette particularité provient de la labilité des synapses qui servent à marquer l'empreinte. On peut donc penser que le programme génétique fournit un moyen d'entretenir automatiquement l'ensemble des traces : on peut par exemple imaginer que le rêve, qui met en jeu un grand nombre de circuits nerveux et qui est une fonction périodique présente chez tous les organismes supérieurs, est l'un de ces moyens, du moins pour certains types de mémoires.
L'apprentissage apparaîtrait ainsi comme la mise en place d'un schéma particulière de l'organisation du réseau; cette trace est en grande partie délocalisée puisqu'elle peut se trouver distribuée sur un très grand nombre de synapses; on peut donc assez bien concevoir qu'elle soit stable par ablation d'une partie de la structure. D'autre part, comme le simple fait d'introduire un message à l'entrée du système fait fonctionner l'organisation du réseau acquise au cours de l'apprentissage et celle-là seulement (puisqu'il y a eu dégénérescence des autres possibles...), il s'agit en fait d'une véritable mémoire (c'est-à-dire d'une donnée non seulement apprise mais susceptible d'être rappelée).
L'évolution synaptique est réglée par la fonction Δ, et il est postulé que c'est le soma postsynaptique qui impose de manière rétrograde, l'évolution, en fonction du message total afférent.(3) Cela entraîne, de façon intuitivement évidente, les propriétés associatives de la mémoire; en effet, deux messages provenant de lieux différents du système et arrivant à un même neurone sont mutuellement en interaction, car la stabilisation des synapse qui les transmettent dépend du message de chacune. Ainsi, dans le réflexe conditionné par exemple, on peut penser que c'est la connectivité acquise au cours de l'expérience individuelle qui produit les associations observées, et l'aspect géographique de l'inconscient freudien paraît de même clairement exprimé... D'autre part, il est clair que l'on n'a aucunement besoin de supposer des synthèses spécifiques correspondant aux objets à mémoriser.
De la structure du système nerveux au comportement
Le système nerveux est aussi et surtout un organe qui commande l'homéostasie de l'être vivant tout entier : il est apparu nécessaire d'étudier un mode formel du couplage entre la réalisation du programme neuronique et le comportement individuel. Pour cela, nous avons dégagé trois applications (au sens mathématique) essentielles qui représentent l'évolution de la structure, du fonctionnement interne et du comportement en fonction de l'apprentissage, considéré comme une trajectoire dans l'environnement. Remarquons que toute interaction, même passive, intervient dans l'évolution du système biologique, ce qui entraîne l'apparence "instructive" de l'environnement. On constate alors que l'organisme biologique formel possède une propriété inattendue : les trois applications dépendantes de l'apprentissage, structure, fonctionnement et comportement, ne peuvent, à partir d'un certain degré de complexité du réseau, être obtenues l'une à partir de l'autre. Cela signifie par exemple que deux structures identiques peuvent conduire à des comportements différents (et réciproquement). Il est donc nécessaire, si l'on veut décrire un organisme biologique complexe, de donner une représentation des trois niveaux considérés et non pas d'un seul, comme c'est généralement le cas ("structuralisme", "fonctionnalisme" ou "behaviourisme").
Un exemple très simple : la jonction neuromusculaire
Avant d'en arriver à la description de partiess complexes du système nerveux central, comme le cervelet, nous nous sommes attachés à l'appliquer au cas du système très simple de la jonction neuromusculaire (voir la figure).
![]()
Développement de la jonction neuromusculaire (1974; d'autres modèles sont possibles, en particulier tenant compte de la diffusibilité du récepteur de l'acétylcholine, et de son potentiel d'agrégation; le processus général du regroupement peut facilement se lier au fait même de fonctionner, comme on le montre dans le cas de l'agrégation des gènes persistants dans les génomes bactériens). A la fin de l'embryogenèse le motoneurone établit plusieurs contacts avec la fibre musculaire. L'arrivée d'un potentiel d'action au niveau de chaque contact libère de l'acétylcholine, qui provoque dans la fibre une onde de dépolarisation qui se propage de manière centrifuge. Le modèle suppose alosrs que, lorsque cette onde arrive aux extrémités de la fibre, elle se réfléchit en un signal de nature différente qui retourne vers la centre de la fibre. La théorie suppose d'autre part que c'est ce signal qui va permettre la stabilisation des contacts : lorsqu'une certaine valeur est atteinte en un point de la fibre, les contacts qui se trouvent dans une petite zone autour de ce point sont stabilisés. Sur le schéma on voit que les zones où naît un signal centrifuge (zone jaune), et les zones où la somme des signaux centripètes issus des deux extrémités de la fibre permet la stabilisation des contacts qui peuvent s'y trouver (zone bleue) se recouvrent pour un signal central (superposition verte du signal jaune et du signal bleu). Pour un contact excentré, la zone de stabilisation n'a aucune raison de recouvrir un contact préexistant. Les fibres nerveuses correspondant aux contacts non stabilisés dégénèrent (tirets). Seule celle du contact central s'établit définitivement.
Les données biologiques sur lesquelles nous nous appuyons sont les suivantes :
— au stade du développement de la jonction neuromusculaire dit des
"fibres exploratrices", le motoneurone établit avec la fibre
musculaire (muscle strié rapide) plusieurs contacts distribués
irrégulièrement, en général sur une grande portion de la fibre
musculaire;
— chez l'adulte, dans un grand nombre de cas, il ne persiste qu'un
seul contact (plaque motrice) au milieu de la fibre;
— le stade intermédiaire de maturation, qui s'accompagne en
particulier de la dégénérescence des contacts éloignés du centre et de
la stabilisation du contact central, est concomitant de l'activité du
nerf et de celle du muscle (motilité embryonnaire);
— la croissance du nerf s'effectuer avec de notables fluctuations,
même dans les lignées hautement consanguines; la position précise de
la plaque motrice peut donc difficilement être programmée
génétiquement.
Afin de rendre compte des faits expérimentaux, les postulats
biologiques suivants, (analogues aux postulats généraux I à V) sont
proposés :
— un contact synaptique peut exister sous au moins trois états,
labile, stable et dégénéré (II). L'étape initiale de reconnaissance au
début du développement peut être décrite par le passage de l'état
néant à l'état labile;
— l'évolution de l'état d'une synapse donnée est gouvernée, de
manière rétrograde, par les signaux reçus par sa membrane
post-synaptique (III). Dans le cas de la contraction musculaire
isométrique, deux types de signaux sont propagés, dans deux canaux
distincts (c'es-à-dire sans interaction entre eux, mais qui peuvent
être géométriquement identiques) : les potentiels d'action
activés au niveau des contacts synaptiques sont propagés par un canal
centrifuge tandis que des signaux centripètes
apparaissent aux extrémités de la fibre, lors de l'arrivée des
potentiels d'action. Une synapse est stabilisée lorsque la somme des
signaux centripètes qui lui parviennent à un instant donné franchit un
seuil critique. Ce seuil est inférieur à la valeur de la somme de deux
signaux centripètes : ainsi, si deux tels signaux parviennent simultanément
sur une synapse, celle-ci se trouve stabilisée;
— l'état connexionnel maximal observé dans la première phase du
développement et le pouvoir évoluteur de la fibre (seuil critique et
signal rétrograde de stabilisation) sont déterminés génétiquement. La
sélection de la synapse centrale requiert une activité particulière du
nerf et du muscle, et constitue donc un processus élémentaire
d'apprentissage (IV).
Ces postulats biologiques permettent une description mathématique à partir de laquelle on peut évaluer semi-quantitativement certains des paramètres postulés et les soumettre ainsi à une vérification expérimentale (4).
Les opérations les plus nobles du cerveau humain sont dues essentiellement à des propriétés innées
En conclusion, il apparaît un parallèle frappant entre la théorie proposée et la théorie darwinienne de l'évolution : dans cette dernière, des fluctuations limitées du patrimoine génétique permettent à l'environnement d'opérer une sélection dans une large population d'individus presque tous identiques, ce qui conduit à une sélection des espèces adaptées de la façon la plus raffinée à leur environnement. Ainsi nous pouvons remarquer (voir JZ Young) que l'information génétique d'une espèce donnée peut déjà être considérée comme représentant une trace de l'environnement, trace qui est l'image de son histoire phylogénétique. D'autre part, dans la théorie de l'apprentissage que nous avons exposée, c'est une épigenèse à partir d'une enveloppe génétique (qui prescrit à un système nerveux certaines règles de fluctuation) qui sélectionne par interaction avec l'environnement une propriété associative stable. Celle-ci est donc aussi une représentation de l'environnement.
Ainsi les opérations les plus nobles du cerveau humain, y compris le langage, sont très probablement dues pour l'essentiel à des propriétés innées; les schèmes généraux et les principales propriétés associativesz sont fournis par un donné héréditaire, et l'acquis n'apparaît que comme une modulation par l'environnement d'une frange fluctuante. Cependant ce schéma, qui pourrait paraître très restrictif, doit être considéré dans toute sa généralité : les réseaux nerveux qui permettent les opérations qui nous paraissent les plus extraordinaires ne sont pas apparus brutalement mais à la suite d'une très longue histoire évolutive. Cette évolution retrace dans les gènes, comme nous l'avons suggéré, une empreinte très riche de l'environnement. L'inné, qui permet le cerveau humain, est déjà une image perfectionnée de l'environnement de l'homme, et c'est une modulation sur un système d'interactions déjà extraordinairement complexe qaui produit les propriétés essentielles de nos apprentissages; et c'est enfin la combinaison entre elles de propriétés complexes qui permet de parvenir à un édifice de complexité qualitativement différente.
Une réflexion à la suite de la réunion du CRSH à Endicott House en 1974 m'a conduit à suivre les travaux de Noam Chomsky, et à les relier à nos travaux sur les mécanismes de stabilisation sélective dans le cerveau au cours de l'apprentissage. Un schéma général de la structure du réseau nerveux comme une collection de modules cherchant à restaurer une homéostasie locale, sans y parvenir, conduit à un processus de balayage du cerveau qui a beaucoup des propriétés de ce qu'on nomme habituellement la conscience.
P Courrège, A Danchin
Apprentissage et changement dans le système nerveux central
Colloque de Cerisy-la-Salle, Change (ed. 10/18) (1975): pp 60-73
A Danchin
Stabilisation fonctionnelle et épigenèse: une approche biologique de
la genèse de l'identité individuelle
In: "L'Identité" (JM Benoist, ed) Grasset (1977): 185-221 ![]()
A Danchin
Spécification épigénétique des réseaux nerveux par stabilisation
fonctionnelle de synapses en développement
In: "Neurobiologie de l'Apprentissage" (R Delacour, ed) Masson
(1978): 198-209
A Danchin
Comment peut-on parler de l'automate cérébral aujourd'hui ?
Revue Philosophique (1980) No 3: 287-304
Depuis le XVIIIe siècle la philosophie a été envahie par un "biologisme" mécaniste. La Mettrie en est l'un des meilleurs exemples. D'où la tentation de parler de l'automate cérébral comme d'un automate (servo-)mécanique. La science du calcul, qui est en révolution depuis le début des années 1930 ne s'accomode pas de la mécanique, même de sa forme élaborée en servo-mécanisme. Il existe en effet un rôle essentiel du lieu du calcul dans son développement. Si, donc, on devait encore parler d'automate cérébral, il serait essentiel de le placer dans le lieu où il apprend sans cesse, dans son environnement. En quelque sorte les approches kantienne doivent s'incarner pour prendre en compte la réalité de l'objet qu'est le cerveau.
A Danchin
Le pilote fantôme (le cerveau et la conscience)
Le Débat (1982) 21: 123-130
A Danchin
Les bases cérébrales du langage
Le Débat (1987) 47: 158-171
A Danchin
Biological foundations of language: a comment on Noam Chomsky's
approach of syntactic structures
In: Noam Chomsky: Consensus and Controversy. (Ed. Drs S& C
Modgil) (1987) pp 29-39
A Danchin
Biological foundations of language: a reply to Dr. Marshall
In: Noam Chomsky: Consensus and Controversy. (Ed. S& C
Modgil) (1987) pp 50-52
A Danchin
Concept et langue, question de neurologie
Colloque Transcultura, (1988), Louvain-la-Neuve
In: "Connaissance et Réciprocité" (ed. Ciaco), pp 41-56
Complément 2009, le rôle du démon de Maxwell, sa myopie, et sa créativité
Le texte qui précède date de 35 années. La plupart de ses propositions sont toujours d'actualité, mais il va de soi qu'un certain nombre de concepts se sont considérablement affinés au cours du temps. En particulier j'ai été amené à lentement élaborer la signification de la dernière phrase du paragraphe qui précède. D'où provient donc la différence qualitative entre ce que nous sommes, et les ancêtres que nous avons en commun avec les grands singes anthropomorphes? Le modèle que nous avions proposé, comme la plupart des théories sélectives, ne s'interrogeait pas sur le mécanisme profond de la sélection, et partait du principe de l'existence d'un large ensemble de possibles, pré-existants en tant que possibles. La sélection devenait la réalisation d'un possible au sein d'un immense ensemble. Nous avions là un reste de pensée platonicienne, qui nous permettait de voir l'évolution un peu comme une sorte de révélation photographique d'archétypes. Cela ne pouvait être satisfaisant.
La discussion avec Noam Chomsky, via mon activité de coordinateur du Centre Royaumont pour une Science de l'Homme, me permettait dès 1974-1975 de comprendre qu'au delà de la contrainte génétique des structures syntaxiques — ce qui est certainement une excellente conjecture — le langage permet tout autre chose. Et ce qu'il apporte provient d'une particularité très originale, dont j'ai peu à peu compris qu'il s'agit de la récursivité. Le langage permet de parler de soi-même. Et cela ouvre un champ totalement nouveau, celui de la création, de novo, plutôt que de la révélation d'un possible préexistant. Nous n'avions conçu notre modèle qu'en termes géométriques et temporels, en le fondant sur les représentation classiques du monde au moyen d'équations aux dérivées partielles. Nous restions implicitement fondamentalement liés aux quatre catégories acceptées du Réel, masse, énergie, espace et temps. Même l'information n'était alors conçue — du moins à notre connaissance — que comme une particularité de la physique statistique, celle des grands ensembles.
Il m'apparaissait pourtant un caractère central de la propagation de
la vie : les enfants ne sont jamais des répliques des parents. Et
au cours d'un séminaire organisé par Jean-Marie
Benoist (disparu en 1990) pour le séminaire annuel de Claude
Lévi-Strauss, je développai cette idée, fondée sur l'apparente
nécessité de la production toujours renouvelée d'innovations absolues
par la vie, dans une conférence intitulée "Stabilisation
fonctionnelle et épigenèse : une approche biologique de la
genèse de l'identité individuelle" que je terminai par la
célèbre phrase de Rimbaud: "Je, est un Autre", pour exprimer ce
pouvoir créateur ![]() .
Cette réflexion sur la création me fit peu à peu me tourner vers
l'analyse des suites de symboles dans un alphabet fini. Cela
conduisait tout naturellement à l'étude du programme génétique
lui-même, qui n'est autre qu'un cas particulièrement intéressant de
telles suites, et à me lancer dans la génomique.
.
Cette réflexion sur la création me fit peu à peu me tourner vers
l'analyse des suites de symboles dans un alphabet fini. Cela
conduisait tout naturellement à l'étude du programme génétique
lui-même, qui n'est autre qu'un cas particulièrement intéressant de
telles suites, et à me lancer dans la génomique.
Le résultat de ces études est paradoxal : alors que pour beaucoup, le cerveau devait ressembler à un ordinateur (et l'idée même de la construction des ordinateurs a longtemps été motivée par l'idée du cerveau électronique) il semble de plus en plus qu'il n'en soit rien, alors que, bien au contraire, il apparaît de plus en plus raisonnable de considérer la cellule (et les organismes plus complexes) comme des ordinateurs faisant des ordinateurs.
Un point commun reste cependant : dans les deux cas une catégorie centrale, distincte des quatre catégories habituelles utilisées pour rendre compte du Réel, information, se trouve au centre de la réflexion. L'étude approfondie des organismes vivants, motivée par l'idée de reconstruire la cellule pour délimiter ce que nous comprenons déjà, et ce que nous ne comprenons pas encore de la vie, mène à comprendre la façon dont l'information est associée à la vie. En bref, les organismes vivants se comportent comme des pièges à information.
1. P. Courrège (1965) Un modèle mathématique des structures élémentaires de parenté. L'Homme 5: 248-290. (retour au texte)
2. J.-P. Changeux (1970) L'inné et l'acquis dans la structure du cerveau. La Recherche 1: 213-221. (retour au texte)
3. C'est en 2009, 35 ans après cette prédiction, que des expériences probantes sont venues conforter cette conjecture. (retour au texte)
4. JP Changeux et A Danchin ont présenté une interprétation biochimique de ce modèle mathématique dans Nature, 264, 705, en 1976 (cf La Recherche, 76, mars 1977), (retour au texte)