Santiago RAMON Y CAJAL
L'innato e l'acquisito : una teoria selettiva dell'apprendimento
Antoine Danchin
Pubblicato su La Neurobiologia Newton Compton
15; 268-276
J.P. Changeux e A. Danchin hanno presentato una interpretazione
biochimica del loro modello matematico su Nature. 264, 705
(1976).
Dopo più di un secolo di ricerche di neurobiologia è ormai evidente che le cellule nervose (neuroni) e più in particolare i contatti che queste cellule stabiliscono fra di loro, o sinapsi, hanno un ruolo fondamentale nel trattamento dell'informazione che, sotto forma di impulsi elettrochimici, percorre le reti nervose.
Nel 1985, nei suoi Lineamenti di una psicologia scientifica, Freud cercò di spiegare il fenomeno della memoria con le conoscenze biologiche del suo tempo. Egli pensò che i neuroni dovessero venire impressionati senza però essere alterati e ciò lo condusse a rifiutare la distinzione tra neuroni della percezione e neuroni del ricordo. Immaginò allora che fosse il cammino tracciato attraverso zone di contatto preesistenti (che noi chiamiamo sinapsi) a permettere la persistenza della memoria: essa sarebbe allora rappresentata dalla differenza tra l'insieme dei percorsi stabiliti ereditariamente e quelli effettivamente acquisity nel corso dell'esperienza individuale; per Freud il principio con cui opera questa selezione è la pipetizione, che permette di tracciare il percorso che costituisce la memoria.
Dopo di allora, numerose osservazionin hanno permesso di precisare queste idee.
Una nuova teoria dell'apprendimento nelle reti nervosi
Per descrivere la capacità del sistema nervoso individuale è opportuno distinguere tre livelli: la struttura del sistema cioè la sua organizzazione spaziale, coi differenti tipi di connessioni; il suo funzionamento, ovvero le regole che definiscono la produzione et l'integrazione dell'attività elettrochimica della rete nervosa, e infine il suo comportamento, cioè il modo in cui un individuo controlla, coll'aiuto del suo sistema nervoso, la sua interazione coll'ambiente.
L'osservazione del sistema nervoso centrale e delle sue proprietà mostra a questi tre livelli, un programma genetico particolarmente rigido; la rigidità delle strutture è ben messa in evidenza dall'esistenza di mutanti che costituiscono le varianti ereditarie del tipo anatomico originale (a livello dell'organizzazione globale, di una classe di neuroni, di una classe di sinapsi...). Allo stesso modo le regole che prescrivono la specificità delle connessioni, sono stabilite con una grande precizione, come le regole che governano la genesi dei potenziali d'azione (impulso nervoso) e il lore ritmo di attività in funzione dello sviluppo dell'individuo. infine, come consenguenza, il comportamento stesso è in grande misura geneticamente determinato.
Tuttavia,Tuttavia, se la rigidità del programma genetico, per i piccoli invertebrati, in generale, appare totale, via via che il sistema nervoso aumenta di complessità, compare una certa fluttuazione: non è che lo schema generale di organizzazione dei tre livelli menzionati ad essere stabilito dal programma. L'evoluzione delle forme di apprendimento più complesse sembra procedere di pari passo coll'aumento di questa fluttuazione: essa compare soprattutto all'inizio dell'ontogenesi, nel corso di un periodo critico, ma permane talora nell'adulto. La stabilità della struttura sembra legata al funzionamento generale del sistema: gli esperimenti di deprivazione sensoriale (dove il giovane animale viene privato dell'interazione col suo ambiente) hanno spesso per effetto degenerazioni profonde dell'organizzazione delle reti nervose.
Queste osservazioni sono alla base di una teoria dell'apprendimento nelle reti nervose, i cui fondamenti sono appena stati pubblicati da J.P. Changeux, P. Courrège, A Danchin. Sitratta di una teoria dell'epigenesi delle reti nervose.
L'epigenesi è la formazione di un'organizzazione, che avviene nel corso dello sviluppo dell'organismo vivente, in seguito alle sue interazioni coll'ambiente. L'apprendimento è considerato come l'acquisizione, nel corso di queste interazioni, di una proprietà associativa ben definita e stabile e viene discusso in termini di epigenesi. La memoria non è che apprendimento suscettibile di essere ricordato (si tratta in questo caso della memoria a lungo termine).
Il modello è stato costruito seguendo il metodo assiomatico, cioè la definizione e lo studio del modello matematico sono accuratamente distinti dal loro significato biologico. Il pensiero biologico non può interferire con il rigore e la coerenza interna del modello matematico e al tempo stesso i problemi posti dalla biologia non possono essere deformati dalla rigidità o dalla schematizzazione grossolana del modello. Sono degli enunciati particolari, chiamati interpretazioni, che permettono di mettere in relazione la biologia colla formalizzazione matematica. In questo senso questo metodo, che è fondato su postulati biologici interpretati da assiomi matematici, è totalmente opposto al metodo abituale che consiste nell'esibire un oggetto matematico le cui proprietà vengono descritte in termini pseudobiologici, e nell'assimilare poi, in seguito a un ragionamento analogico, il modello alla realtà sperimentale.
Postulati biologici e assiomi matematici
I postulati biologici che ci sembrano pertinenti e sufficienti per una descrizione delle caratteristiche fondamentali dell'apprendimento, della memoria, e delle proprietà pià evolute del sistema nervoso centrale dei vertebrati, sono i seguenti:
— I. I neuroni sono i soli responsabili del trattamento dell'informazione che ha luogo nel sistema nervoso: viene considerato solo il carattere discontinuo degli impulsi che essi creano, integrano e propagano.
— II. I contatti tra i neuroni o sinapsi sono gli intermediaridi tutti i trasferimenti di informazione attraverso il sistema. Queste sinapsi possono esistere almeno in tre stati (plasticità sinaptica): labile (L), stabile (S) e degenerato (D); solo L e S trasmettono l'impulso nervoso. Le transizioni permesse sono
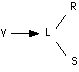
— III. L'evoluzione dello stato di ciascuna sinapsi è regolato dall'insieme degli impulsi che arrivano sul neurone postsinaptico in un intervallo di tempo di lunguezza determinata (si dice che il soma neuronico ha un potere evolutore).
— IV. Il numero massimo di connessioni, le tappe dello sviluppo della rete e il potere integratore o evolutore di ogni soma neuronico, sono determinati in modo rigoroso dal programma generico: è questo l'inviluppo genetico della rete.
— V. L'apprendimento è considerato come un'attitudine acquisita, associata alla variabilità dell'organizzazione delle connessioni del sistema.
Ciò che è innato è dunque il vincolo genetico mentre ciò cheviene acquisito è una proprietà associativa stabile. L'apprendimento è il processo attraverso il quale una certa classe di interazioni con l'ambiente permette la messa a punto di questa proprietà associativa. Secondo la teoria proposta, quest'ultima si troverebbe inscritta nella struttura (e nel funzionamento) della rete nervosa, in una particolare forma di organizzazione della rete. Questa forma risulta in generale dalla selezione di alcuni cammini stabili tra molte possibilità fornite dalla comparsa di un gran numero di sinapsi labili nel corso dello sviluppo. Ci si possono immaginare i circuiti nervosi come una immensa rete a tre dimensioni i cui nodi sono connessi gli uni agli altri in una enorme quantità di giunzioni: in seguito all'interazione coll'ambiente una parte delle connessioni viene tagliata e l'immagine particolare dell'ambiente rimane scolpita nella rete.
Le realizzazioni di un programma neuronico
Dal punto di vista formale, viene studiato un oggetto matematicochiamato programma neuronico R = (C, Σ, Θ, n, Φ, Δ) dove, (C, Σ) è un grafo particolare in cui possono essere distinte delle entrate e delle uscite che vengono interpretate come massima struttura connessionale possibile; Θ rappresenta per ogni connessione il ritardo di trasvisione dell'impulso nervoso tra il suo punto di emergenza e il neurone successivo; Φ rappresenta il potere di integrazione che permette a ogni neurone di inviare un impulso nervoso in funzione del messaggio totale che riceve durante l'intervallo di n; Δ infine rappresenta il potere di evoluzione di ogni neurone su ciascuna delle sue sinapsi afferenti, in funzione del programma genetico (della crescita viene tenuto conto supponendo che una connessione si trovi in uno stato nullo N prima di apparire in uno stato labile) e del messaggio totale afferente durante l'intervallo n.
Oggetto di questa teoria è lo studio delle proprietà delle realizzazioni di questo programma neuronico in funzione di un multimessaggio imposto sulle entrate del grafo neuronico, che rappresenta la fase « istruttiva » dell'apprendimento. Una classe di programmi neuronici può acquistare una competenza T-T', cioè in un periodo di tempo finito, dei messaggi di un dato tipo T sono trasformati in messaggi di tipo T' stabili all'uscita: in generale una competenza di questo tipo non rimane stabile che alla condizione che un funzionamento minimo venga imposto all'entrata della rete; d'altra parte si osserva che nel periodo di acquisizione della competenza T-T', uno scarso funzionamento provoca ampie degenerazioni (cià si accorda bene cogli esperimenti di deprivazione sensitiva). Così l'espressione del programma genetico non può in certi casi svilupparsi correttamente che a condizione che la rete nervosa sia mantenuta in funzione. È qui che interviene in modo privilegiato l'ambiente e viene lasciata l'improntadi cià che è appreso. Questa particolarità dipende dalla labilità delle sinapsi che servono a marcare l'impronta. Si può dunque pensare che il programma genetico fornisca un mezzo perconservare automaticamente l'insieme delle tracce: si può adesempio immaginare che il sogno, che mette in azione un gran numero di circuiti nervosi e che è una funzione periodica presente in tutti gli organismi superiori, sia uno di questi mezzi, valido almeno per un certo tipo di memorie.
L'apprendimento apparirebbe cosà la messa a punto di uno schema particolare di organizzazione della rete; questa traccia è in gran parte delocalizzata poiché può trovarsi distribuita su un gran numero di sinapsi; si può dunque pensare che essa rimanga stabile dopo l'ablazione di una parte della struttura. D'altra parte, poiché il semplice fatto di introdurre un messaggio all'entrata del sistema fa funzionare l'organizzazione della rete acquisita nel corso dell'apprendimento ed essa soltanto (poiché sono degenerate le altre possibili), si tratta infatti di una vera e propria memoria (cioè di un dato non solamente appreso maanche suscettibile di essere ricordato).
L'evoluzione sinaptica è regolata dalla funzione Δ ed è postulato che sia il soma postsinaptico che impone, in modo retrogrado, l'evoluzione, in funzione della totalità dei messaggi afferenti. Ciò provoca, in modo intuitivamente evidente, le proprietà associative della memoria; infatti due messaggi provenienti da luoghi differenti del sistema, che giungono a uno stesso neurone, interagiscono reciprocamente poiché la stabilizzazione delle sinapsi cheli trasmettono dipende dal messaggio di ciascuna. Così, nel riflesso condizionato ad esempio, si può pensare che sia la connessione acquisita nel corso dell'esperienza individuale che produce le associazioni osservate, e anche l'aspetto geografico dell'inconscio freudiano sembra interpretabile in modo analogo... D'altra parte è chiaro che non si ha alcun bisogno di supporre l'esistenza di sintesi specifiche in corrispondenza degli oggetti da memorizzare.
Dalla struttura del sistema nervoso al comportamento
Il sistema nervoso è anche, e soprattutto un organo che dirige l'omeostasi dell'essere vivente in tutta la sua completezza: ci è sembrato necessario studiare un tipo di accoppiamento formale tra la realizzazione del programma neuronico e il comportamento individuale. Abbiamo allora considerato tre applicazioni (in senso matematico) essenziali che rappresentano l'evoluzione della struttura, del funzionamento interno e del comportamento, in funzione dell'apprendimento, considerato come una traiettoria nell'ambiente. Si osserva allora come l'organismo biologico formale possegga una proprietà inattesa: le tre applicazioni dipendenti dell'apprendimento, struttura, funzionamento e comportamento, non possono, oltre un certo grado di complessità essere ottenute l'una a partire dall'altra. Ciò significa ad esempio che due strutture identiche possono condurre a comportamenti differenti (e vice versa). È dunque necessario, se si vuole descrivere un organismo biologico complesso, dare una rappresentazione ditutti e tre i livelli considerati e non di uno solo, come accade disolito (« strutturalismo », « funzionalismo » o « behaviourismo »).
Un esempio molto semplice: la giunzione neuromuscolare
Prima di giungere alla descrizione di parti complesse del sistema nervoso centrale, come il cervelietto, vogliamo considerare il caso molto semplice del sistema costituito dalla giunzione neuromuscolare(vedi figura).
I dati biologici sui quali ci basiamo sono i seguenti:
a. allo stadio dello sviluppo della giunzione neuromuscolare detto delle « fibre esploratrici », il motoneurone stabilisce con la fibra muscolare (muscolo striato rapido) molti contatti distribuiti in modo irregolare, generalmente su di un'ampia porzione dellafibra muscolare;
b. nell'adulto, in un gran numero di casi non rimane che unsolo contatto (placca motrice) in mezzo alla fibra;
c. lo stadio intermedio di maturazione, che è accompagnato dalla degenerazione dei contatti lontani dal centro e dalla stabilizzazione del contatto centrale, è concomitante con l'attività del nervo e con quella del muscolo (movimenti embrionali);
d. nella crescita del nervo intervengono notevoli variazioni, anche all'interno di linee notevolmente consanguinee; la posizione precisa della placca motrice può dunque difficilmente essere programmata geneticamente.
![]()
Sviluppo della giunzione neuromuscolare. Alla fine dell'embriogenesi il motoneurone stabilisce numerosi contatti con la fibra muscolare. L'arrivo di un potenziale d'azione a livello di ogni contatto libera dell'acetilcolina, che provoca nella fibra un'onda di depolarizzazione che si propaga in modo centrifugo. Il modello presuppone allora che, quando quest'onda giunge all'estremità della fibra essa si rifletta in un segnale di natura differente che torna verso il centro della fibra. La teoria prevede inoltre che sia questo segnale che permette la stabilizzazione dei contatti: quando viene raggiunto un certo valore soglia in un punto della fibra, i contatti che si trovano in una piccola zona intorno alla fibra vengono stabilizzati. Sullo schema si vede come le zone dove nasce un segnale centrifugo (tratteggiato) e le zone in cui la somma dei segnali centripeti provenienti dalle due estremità della fibra permettono la stabilizzazione dei contatti che vi si trovano (punteggiato) si sovrappongano nel caso di un contatto centrale. Nel caso di un contatto non centrato, la zona di stabilizzazione non ha alcun motivo per ricoprire un contatto preesistente. Le fibre nervose corrispondenti ai contatti non stabilizzati degenerano (lineette). Solo quella del contatto centrale si stabilisce definitivamente.
Per rendere conto dei risultati sperimentali vengono proposti i seguenti postulati biologici (analoghi ai postulati generali da I a V).
a. Un contatto sinaptico può trovarsi per lo meno in trestati: labile, stabile e degenerato (II). La tappa iniziale di riconoscimento d'inizio dello sviluppo può essere descritta come il passaggio dallo stato nullo a quello labile.
b. L'evoluzione dello stato di una data sinapsi è governatoin modo retrogrado, dai segnali ricevuti dalla sua membrana postsinaptica (III). Nel caso della contrazione muscolare isometrica, vengono propagati due tipi di segnali, attraverso due canali distinti (cioè senza interazione reciproca), ma che possono essere geometricamente identici): i potenziali d'azione generati a livellodel contatto sinaptico vengono propagati attraverso un canale centrifugo mentre i segnali centripeti appaiono alle estremità della fibra, quando giunge il potenziale d'azione. Una sinapsi viene stabilizzata quando la somma dei potenziali d'azione che la raggiungono raggiunge una soglia critica. Questa soglia è inferiore al valore della somma di due segnali centripeti: cosà se due segnali di questo tipo raggiungono simultaneamente una sinapsi essa viene stabilizzata.
c. Lo stato con numero di connessioni massimo osservato nella prima fase dello sviluppo e il potere evolutore della fibra (soglia critica e segnale retrogrado di stabilizzazione) sono determinati geneticamente. La selezione della sinapsi centrale richiede un'attività particolare del nervo e del muscolo e costituisce dunque un elementare processo di apprendimento (IV).
Questi postulati biologici permettono una descrizione matematica con la quale si possono valutare semiquantitativamente alcunidei parametri postulati e sottoporli cosà a una verifica sperimentale'.
Le più nobili operazioni del cervello umano sono dovute essenzialmente a delle proprietà innate
In conclusione si delinea un sorprendente parallelismo tra la teoria proposta e quella darwiniana dell'evoluzione: in quest'ultima variazioni limitate del patrimonio genetico permettono all'ambiente di operare una selezione su una larga popolazione di individui quasi tutti identici che ha come risultato l'evoluzione di specie adattate nel modo pià raffinato al loro ambiente. Possiamo dunque notare (vedi J.Z. Young) che l'informazione genetica di una data specie può già essere considerata la rappresentazione di una traccia dell'ambiente, traccia che è l'immagine della sua storia filogenetica. D'altra parte, nella teoria dell'apprendimento che abbiamo esposto è l'epigenesi che parte da un programma genetico (che prescrive al sistema nervoso alcune regole di fluttuazione) che seleziona interagendo coll'ambiente una proprietà associativa stabile. Anche questa è dunque una rappresentazione dell'ambiente.
Le più nobili operazioni del cervello umano, ivi compreso il linguaggio, sono molto probabilmente dovute per l'essenziale aproprietà innate; gli schemi generali del programma ereditario e ciò che viene acquisito non compare che come modulazione indotta dall'ambiente, delle fluttuazioni del programma. Questo schema che potrebbe apparire troppo restrittivo deve tuttavia essere considerato in tutta la sua generalità : le reti nervose che permettono le operazioni che ci appaiono cosi straordinarie non sono comparse improvvisamente, ma sono il risultato di una lunga storia evolutiva. Questa evoluzione lascia nei geni, come abbiamo suggerito, una impronta molto dettagliata dell'ambiente. Le strutture innate che permettono la realizzazione del cervello umano sono già una immagine perfezionata del suo ambiente. È la modulazione di un sistema di interazioni già straordinariamente complesso a fare da base alle proprietà essenziali del nostro apprendimento ed è infine la combinazione tra loro di proprietà complesse che permette l'edificazione di un sistema la cui complessità è qualitativamente differente.